
ΦΑΚΕΛΛΟΣ "ΙΟΛΟΓΙΑ": Η Υποθετική Μετατροπή του RNA σε cDNA


Μετάφραση: Απολλόδωρος
19 Φεβρουαρίου 2022 | Mike Stone | Διαβάστε το εδώ

Προκειμένου να δημιουργηθεί ένα "ιογενές" γονιδίωμα, υπάρχουν πολλά βήματα και διαφορετικές διαδικασίες από τις οποίες πρέπει πρώτα να περάσει το δείγμα για να προετοιμαστεί για την αλληλούχιση. Κανονικά το δείγμα υποβάλλεται σε τοξική καλλιέργεια κυττάρων προκειμένου να αναπτυχθεί αρκετός "ιός" ώστε να χρησιμοποιηθεί το μη καθαρισμένο υπερκείμενο της καλλιέργειας για την αλληλούχιση. Ορισμένες φορές η αλληλούχιση γίνεται απευθείας στα μη καθαρισμένα υγρά από άρρωστους ασθενείς, όπως έγινε στην περίπτωση του "SARS-COV-2". Όποια μέθοδος και αν χρησιμοποιείται, το δείγμα πρέπει να διασπαστεί για να εξαχθεί το RNA που απαιτείται για την αλληλούχιση. Ωστόσο, υπάρχει ένα μικρό πρόβλημα. Λέγεται ότι συνήθως δεν μένει αρκετό υλικό στο τέλος της εξαγωγής του RNA για την αλληλουχία ενός γονιδιώματος, οπότε είναι ανάγκη να δημιουργηθεί περισσότερο. Προκειμένου να δημιουργηθεί το απαιτούμενο υλικό, πρέπει να χρησιμοποιηθεί PCR για ενίσχυση. Δυστυχώς, υποστηρίζεται ότι η PCR μπορεί να χρησιμοποιηθεί μόνο για την ενίσχυση του DNA και όχι του RNA. Έτσι, ο μόνος τρόπος για να λειτουργήσει αυτό είναι να χρησιμοποιηθεί μια διαδικασία γνωστή ως Αλυσιδωτή Αντίδραση Πολυμεράσης με Αντίστροφη Μεταγραφή (RT-PCR) για να μετατραπεί το RNA σε DNA. Από την RT-PCR, το RNA μετατρέπεται σε αυτό που ονομάζεται συμπληρωματικό DNA ή cDNA με τη χρήση ενός ενζύμου γνωστού ως αντίστροφη μεταγραφάση, εξ ου και το RT στην RT-PCR. Αυτό γίνεται προκειμένου να δημιουργηθεί αυτό που ονομάζεται βιβλιοθήκη αλληλούχισης:
Βιβλιοθήκη αλληλούχισης: Τι είναι;
Βιβλιοθήκες αλληλούχισης DNA και RNA
"Μια βιβλιοθήκη αλληλούχισης μπορεί να δημιουργηθεί ξεκινώντας από γονιδιωματικό DNA ή από RNA. Η ροή εργασίας για την προετοιμασία μιας βιβλιοθήκης αλληλούχισης DNA αποτελείται από τρία βασικά βήματα:
Κατακερματισμός και διαστασιολόγηση του νουκλεϊκού οξέος (DNA ή RNA) για τη λήψη θραυσμάτων προκαθορισμένου μήκους.
Προσάρτηση των προσαρμογέων (adapters) στα άκρα των θραυσμάτων
Ποσοτικοποίηση της βιβλιοθήκης
Σε κάθε βιβλιοθήκη αλληλούχισης RNA υπάρχει ένα πρόσθετο βήμα: η μετατροπή του RNA σε cDNA. Το βήμα της κατακερματισμού μπορεί να γίνει πριν ή μετά τη σύνθεση του cDNA".


Η βιβλιοθήκη αλληλούχισης είναι ουσιαστικά μια δεξαμενή θραυσμάτων DNA με προσαρτητές συνδεδεμένους. Η εργασία με RNA, όπως κατά την αλληλούχιση ενός "ιού", απαιτεί το πρόσθετο βήμα της μετατροπής cDNA πριν από την προετοιμασία αυτής της βιβλιοθήκης. Αυτή η μετατροπή σε cDNA γίνεται για μερικούς λόγους:
Το RNA θεωρείται πιο ασταθές από το DNA.
Η ενίσχυση με PCR λειτουργεί μόνο σε DNA.
Τα περισσότερα, αν όχι όλα, τα πρωτόκολλα αλληλούχισης είναι σχεδιασμένα για DNA.
Τι ακριβώς είναι λοιπόν το cDNA και πώς μπορεί κανείς να το δημιουργήσει; Η θεωρία για την εξήγηση του cDNA στηρίζεται σε μεγάλο βαθμό στην απάτη περί "ιού", καθώς ο μόνος τρόπος για να υπάρξει cDNA είναι είτε μέσω της χρήσης της αντίστροφης μεταγραφάσης, ενός ενζύμου που δημιουργείται μέσω των "φυσικών" διαδικασιών των "ρετροϊών" καθώς καταλαμβάνουν κύτταρα ξενιστές (δηλ. μυθοπλασία), είτε ως συνθετικά κατασκευασμένο DNA σε ένα εργαστήριο με τη χρήση της συνθετικά δημιουργημένης αντίστροφης μεταγραφάσης (δηλ. μυθοπλασία). Με άλλα λόγια, ο μόνος τρόπος ύπαρξης του cDNA είναι μέσω της χρήσης ενός φανταστικού ενζύμου από έναν φανταστικό "ιό" που δημιουργήθηκε είτε από κυτταρική καλλιέργεια σε εργαστήριο είτε με συνθετικά μέσα μέσω RT-PCR:
cDNA έναντι Γονιδιωματικού DNA
"Αρχικά, παρατηρήθηκε ότι το gDNA διαβάζεται πάντα και μεταγράφεται σε mRNA, το οποίο καθοδηγεί το σχηματισμό πρωτεϊνών και στη συνέχεια απορρίπτεται. Η αντίληψη ότι η πληροφορία μπορεί πάντα να ρέει από το DNA στο RNA στην πρωτεΐνη αναφέρθηκε κάπως χαριτολογώντας ως το Κεντρικό Δόγμα της μοριακής βιολογίας. Η ονομασία αυτή προκάλεσε τους επιστήμονες να βρουν εξαιρέσεις σε αυτόν τον κανόνα.
Οι ιολόγοι βρήκαν τελικά μια τέτοια εξαίρεση. Ανακαλύφθηκε ότι οι ρετροϊοί διαθέτουν μηχανισμούς "αντίστροφης μεταγραφής". Αυτό σημαίνει ότι μπορούν να παίρνουν αλυσίδες RNA και να παράγουν αλυσίδες DNA από αυτές. Με αυτόν τον τρόπο, κατά την αντίστροφη μεταγραφή, η πληροφορία ρέει αντίστροφα από το RNA πίσω στο DNA. Το DNA που προκύπτει από αυτή τη διαδικασία ονομάζεται συμπληρωματικό DNA (cDNA). cDNA παράγεται είτε από ορισμένους ιούς είτε συντίθεται στα εργαστήρια.
Το cDNA μπορεί να περιγραφεί ως gDNA χωρίς όλες τις απαραίτητες μη κωδικοποιητικές περιοχές, και έτσι παίρνει το όνομά του ως συμπληρωματικό DNA".
"Όταν οι επιστήμονες χρησιμοποιούν ιικά ένζυμα για να παρασκευάσουν cDNA από RNA που απομονώθηκε από τα κύτταρα και τους ιστούς που μελετούν, αυτό δεν περιέχει ιντρόνια λόγω του ότι έχει συγκολληθεί στο mRNA. Το cDNA δεν περιέχει επίσης κανένα άλλο gDNA που δεν κωδικοποιεί άμεσα μια πρωτεΐνη (αναφέρεται ως μη κωδικοποιητικό DNA)."
"Για να απομονωθεί cDNA, πρέπει πρώτα να απομονωθεί το RNA ενός οργανισμού. Στη συνέχεια, χρησιμοποιώντας ένα ένζυμο αντίστροφης μεταγραφάσης, μπορεί να παραχθεί cDNA. Αυτή είναι η διαδικασία που χρησιμοποιούν οι ρετροϊοί για να ενσωματωθούν στα κύτταρα του ξενιστή τους. Οι ρετροϊοί, όπως ο ιός της ανοσοανεπάρκειας των πιθήκων (SIV) και ο ιός της μυελοβλάστης των πτηνών (AMV), χρησιμοποιούν το cDNA τους για να παράγουν mRNA στον ξενιστή, οδηγώντας στην παραγωγή ιικών πρωτεϊνών. Αυτό είναι δυνατό επειδή οι ρετροϊοί χρησιμοποιούν RNA ως γονιδιωματικό τους υλικό αντί για DNA και αυτό μεταγράφεται αντίστροφα στο cDNA, το οποίο στη συνέχεια υφίσταται κανονική μεταγραφή και οδηγεί στην ιική πρωτεΐνη στον ξενιστή".
https://www.biochain.com/blog/cdna-vs-genomic-dna/
Όπως γίνεται αντιληπτό, το RNA υποβάλλεται στη μαγική διαδικασία της RT-PCR χρησιμοποιώντας την αντίστροφη μεταγραφάση που έχει κατασκευαστεί από έναν "ιό" και έχει δημιουργηθεί σε ένα εργαστήριο. Για να το θέσουμε σε προοπτική, ας αναλύσουμε λίγο αυτή τη διαδικασία χρησιμοποιώντας το γονιδίωμα "SARS-COV-2" ως παράδειγμα. Προκειμένου να δημιουργηθεί το γονιδίωμα για τον "SARS-COV-2", το μη καθαρισμένο BALF ενός ασθενούς υποβλήθηκε σε βαθιά μετα-τρανσκριπτομική αλληλουχία. Το RNA εξήχθη από αυτό το μη καθαρισμένο BALF και κατασκευάστηκε μια βιβλιοθήκη για την αναζήτηση "αιτιολογικών παραγόντων". Από τα 384.096 contigs που συναρμολογήθηκαν από το Megahit9, διαπίστωσαν ότι το μακρύτερο (30.474 νουκλεοτίδια (nt)) είχε στενή σχέση με έναν "κορονοϊό" που μοιάζει με τον SARS της νυχτερίδας. Χρησιμοποίησαν RT-PCR για να προσδιορίσουν και να επιβεβαιώσουν το γονιδίωμα και το τεριμίνιο του:
"Το ολικό RNA εξήχθη από 200 μl BALF και κατασκευάστηκε μια μετα-τρανσκριπτομική βιβλιοθήκη για αλληλούχιση κατά ζεύγη (150-bp reads) με τη χρήση Illumina MiniSeq, όπως περιγράφηκε προηγουμένως4,6,7,8. Συνολικά, δημιουργήσαμε 56.565.928 αναγνώσεις αλληλουχίας που συναρμολογήθηκαν de novo και ελέγχθηκαν για πιθανούς αιτιολογικούς παράγοντες. Από τα 384.096 contigs που συναρμολογήθηκαν με το Megahit9 , το μεγαλύτερο (30.474 νουκλεοτίδια (nt)) είχε υψηλή αφθονία και ήταν στενά συνδεδεμένο με μια απομόνωση του κορονοϊού (CoV) που μοιάζει με τον SARS της νυχτερίδας - νυχτερίδα SL-CoVZC45 (αριθμός καταχώρησης GenBank MG772933) - που είχε προηγουμένως δειγματοληπτηθεί στην Κίνα, με νουκλεοτιδική ταυτότητα 89,1% (Συμπληρωματικοί πίνακες 1, 2). Η αλληλουχία του γονιδιώματος αυτού του ιού, καθώς και οι άκρες του, προσδιορίστηκαν και επιβεβαιώθηκαν με αντίστροφη μεταγραφή PCR (RT-PCR)10 και ταχεία ενίσχυση των άκρων cDNA 5′/3′ (RACE), αντίστοιχα. Το εν λόγω στέλεχος του ιού ονομάστηκε WH-Human 1 coronavirus (WHCV) (και έχει επίσης αναφερθεί ως "2019-nCoV") και η αλληλουχία ολόκληρου του γονιδιώματός του (29.903 nt) έλαβε τον αριθμό πρόσβασης GenBank MN908947".
Κοιτάζοντας την ενότητα "Μέθοδοι", βρίσκουμε ακριβώς τι χρησιμοποίησαν για να μετατρέψουν το RNA σε cDNA:
"Το ολικό RNA εξήχθη από το δείγμα BALF χρησιμοποιώντας το κιτ RNeasy Plus Universal Mini (Qiagen) σύμφωνα με τις οδηγίες του κατασκευαστή. Η ποσότητα και η ποιότητα του διαλύματος RNA αξιολογήθηκε με τη χρήση μηχανής Qbit και βιοαναλυτή Agilent 2100 (Agilent Technologies) πριν από την κατασκευή βιβλιοθήκης και την αλληλούχιση. Στη συνέχεια κατασκευάστηκε μια βιβλιοθήκη RNA χρησιμοποιώντας το κιτ SMARTer Stranded Total RNA-Seq v.2 (TaKaRa)".
https://www.nature.com/articles/s41586-020-2008-3
Λέει ότι η βιβλιοθήκη δημιουργήθηκε χρησιμοποιώντας το SMARTer Stranded Total RNA-Seq kit v.2 (TaKaRa). Αν πάμε στον κατασκευαστή αυτού του προϊόντος, μπορούμε να βρούμε αυτό το έγγραφο που περιγράφει τη διαδικασία δημιουργίας της βιβλιοθήκης, το οποίο μας δίνει το όνομα της RT που χρησιμοποιείται για τη δημιουργία του cDNA:
"Ο τυχαίος εκκινητής (που αντιπροσωπεύεται ως ο πράσινος εκκινητής N6) επιτρέπει τη δημιουργία cDNA από όλα τα τμήματα RNA του δείγματος, συμπεριλαμβανομένου του rRNA. Όταν η αντίστροφη μεταγραφάση (RT) του SMARTScribe φτάσει στο 5′ άκρο του θραύσματος RNA, η δραστηριότητα τερματικής τρανσφεράσης του ενζύμου προσθέτει μερικά μη υποδειγματικά νουκλεοτίδια στο 3′ άκρο του cDNA (απεικονίζεται ως Xs)".
Διαπιστώνουμε ότι χρησιμοποιούν μια αντίστροφη μεταγραφάση που ονομάζεται SMARTScribe Reverse Transcriptase (RT). Αν αναζητήσουμε το ίδιο το προϊόν, λαμβάνουμε αυτή την περιγραφή που περιγράφει λεπτομερώς τι ακριβώς είναι:
"Η αντίστροφη μεταγραφάση SMARTScribe είναι ένα ένζυμο υψηλής απόδοσης που εκτελεί αμερόληπτη σύνθεση cDNA, επιτρέποντας την ενίσχυση και την κατασκευή βιβλιοθήκης από οποιοδήποτε μεταγράφημα RNA. Το SMARTScribe RT είναι μια τροποποιημένη αντίστροφη μεταγραφάση του ιού της λευχαιμίας του ποντικού Moloney που παράγει cDNA μεγάλου μήκους, πλήρους μήκους (έως 14,7 kb), διατηρώντας παράλληλα τις σχετικές αναλογίες των μεταγραφών του αρχικού δείγματος RNA. Η ιδιόκτητη διαδικασία καθαρισμού μας και τα αυστηρά πρότυπα ελέγχου ποιότητας διασφαλίζουν ότι σχεδόν όλες οι μολυσματικές νουκλεάσες έχουν απομακρυνθεί από το SMARTScribe RT. Έχει διαμορφωθεί ειδικά για χρήση με όλα τα κιτ SMART μας".
https://www.takarabio.com/products/cdna-synthesis/reverse-transcriptases/smartscribe6th
Έτσι, βλέπουμε ότι το RNA από το μη καθαρισμένο BALF υποβλήθηκε σε τροποποιημένη αντίστροφη μεταγραφάση Moloney Murine Leukemia "Virus "Virus. Αν και δεν μπόρεσα να βρω την ακριβή διαδικασία που χρησιμοποιήθηκε για τη δημιουργία αυτής της συγκεκριμένης RT, κρίνοντας από τις περιγραφές πολλών άλλων προϊόντων RT του M-MLV εκεί έξω, πιθανότατα δημιουργήθηκε συνθετικά σε ένα εργαστήριο με "απομόνωση" από E. coli που εκφράζει ένα τμήμα του γονιδίου pol του M-MLV σε ένα πλασμιδιακό γονίδιο. Με άλλα λόγια, πρόκειται για ένα ανασυνδυασμένο γενετικά τροποποιημένο δημιούργημα από πολλαπλές πηγές. Αυτή η RT χρησιμοποιήθηκε για τη δημιουργία του συνθετικού cDNA προκειμένου να αλληλουχηθεί το γονιδίωμα του "SARS-COV-2". Βλέπουμε επίσης ότι η ιδιόκτητη διαδικασία καθαρισμού μπορεί να απομακρύνει σχεδόν όλες τις μολυσματικές νουκλεάσες. Με άλλα λόγια, απομακρύνει τη μόλυνση "στην πραγματικότητα αν και όχι στην πραγματικότητα- πρακτικά- σχεδόν". Διαφορετικά δηλώνεται ως: δεν απομακρύνει όλες τις μολυσματικές ουσίες. Η προσθήκη αντίστροφης μεταγραφάσης που παράγεται από μη καθαρισμένο "ιό" για τη δημιουργία του cDNA είναι μια ακόμη πρόσμειξη που προστίθεται στο δείγμα πριν από την αλληλούχιση.
Για να πιστέψει κανείς στη νομιμότητα της μετατροπής του RNA σε cDNA μέσω της RT-PCR, πρέπει πρώτα να πιστέψει ότι ο "ιός" της λευχαιμίας του ποντικού και ο "ιός" του σαρκώματος των πτηνών (όπως επίσης χρησιμοποιήθηκε) καθαρίστηκαν και απομονώθηκαν. Καθώς είναι πολύ σαφές ότι κανένας "ιός" δεν έχει ποτέ καθαριστεί και απομονωθεί σωστά, αυτό τορπιλίζει αμέσως τη νομιμότητα αυτής της διαδικασίας. Εάν δεν είναι σε θέση να καθαρίσουν και να απομονώσουν έναν "ιό", δεν θα είναι σε θέση να αλληλουχήσουν, να ταυτοποιήσουν και να απομονώσουν οποιαδήποτε γονίδια που λέγεται ότι ανήκουν στην αντίστροφη μεταγραφάση από τον εν λόγω "ιό".
Αν δεχτούμε ότι πράγματι απομόνωσαν τόσο τον MLV όσο και τον ASV, τότε θα πρέπει να πιστέψουμε ότι αυτοί οι "ιοί" αλληλουχήθηκαν με ακρίβεια και αξιοπιστία χωρίς τη χρήση εξελιγμένης τεχνολογίας αλληλούχισης, καθώς η RT ανακαλύφθηκε στις αρχές της δεκαετίας του 1970, δηλαδή πολύ πριν από την τεχνολογική πρόοδο της γονιδιωματικής με την αλληλούχιση Sanger στις αρχές της δεκαετίας του 1980. Ακόμη και σήμερα, η τεχνολογία αλληλούχισης που λέγεται ότι είναι πολύ πιο προηγμένη είναι επιρρεπής σε περιορισμούς, σφάλματα και μη αναπαραγώγιμα αποτελέσματα. Τεντώνει τη φαντασία μας να πιστέψουμε ότι αυτό ήταν δυνατό πριν από 50 χρόνια, όταν δεν είναι ακόμα δυνατό σήμερα.
Αν υποθέσουμε ότι η αλληλουχία RT ήταν κατά κάποιον τρόπο σωστή, τότε πρέπει να πιστέψουμε ότι ήταν σε θέση να αναπαραχθεί συνθετικά σε ένα εργαστήριο συνδυάζοντας το γονίδιο pol του MLV με το E. Coli. Όπως περιγράφεται λεπτομερώς εδώ, ενώ υποστηρίχθηκε ότι αυτή η διαδικασία είχε γίνει, δεν περιγράφεται λεπτομερώς στις αρχικές μελέτες:
"Η αντίστροφη μεταγραφάση (RT) του ιού της λευχαιμίας του ποντικού Moloney (MMLV-RT) είναι το πιο ευρέως χρησιμοποιούμενο ένζυμο για τη σύνθεση cDNA και την ενίσχυση RNA, λόγω της ισχυρής καταλυτικής του δραστηριότητας και της υψηλής πιστότητάς του (Kimmel and Berger 1987- Kievits et al. 1991). Παρόλο που έχει παραχθεί ανασυνδυασμένη MMLV-RT σε Escherichia coli (Tanese et al. 1985- Roth et al. 1985- Kotewicz et al. 1985), οι συνθήκες έκφρασης και καθαρισμού δεν ήταν λεπτομερείς και η στρατηγική καθαρισμού τεσσάρων έως πέντε βημάτων καθιστά αναγκαίο τον σχεδιασμό μιας εναλλακτικής απλής και γρήγορης μεθόδου για την παραγωγή αυτού του ενζύμου σε διαλυτή και ενεργή μορφή".
doi: 10.1007/s10529-009-9977-5.
Η μετατροπή του RNA σε cDNA απαιτεί πολλά άλματα στη λογική προκειμένου να πιστέψει κανείς ότι αυτό που ισχυρίζεται ότι συμβαίνει λαμβάνει πράγματι χώρα. Αυτό που συμβαίνει στο πλαίσιο αυτών των χημικών αντιδράσεων είναι εντελώς μη παρατηρήσιμο. Οι ιστορίες που δημιουργούνται γύρω από αυτά τα πειράματα είναι εντελώς υποθετικές. Η νομιμότητα αυτής της διαδικασίας εξαρτάται από την ύπαρξη ενός αόρατου "ιού" που χρησιμοποιείται για τη δημιουργία του συνθετικού ενζύμου της αντίστροφης μεταγραφάσης προκειμένου να μετατραπεί το RNA σε συνθετικό cDNA, ώστε να δημιουργηθεί ένα θεωρητικό γονιδίωμα για άλλους αόρατους "ιούς". Το να πούμε ότι η πίστη σε αυτή τη διαδικασία απαιτεί μεγάλη τυφλή πίστη είναι μια μετριοπαθής έκφραση..
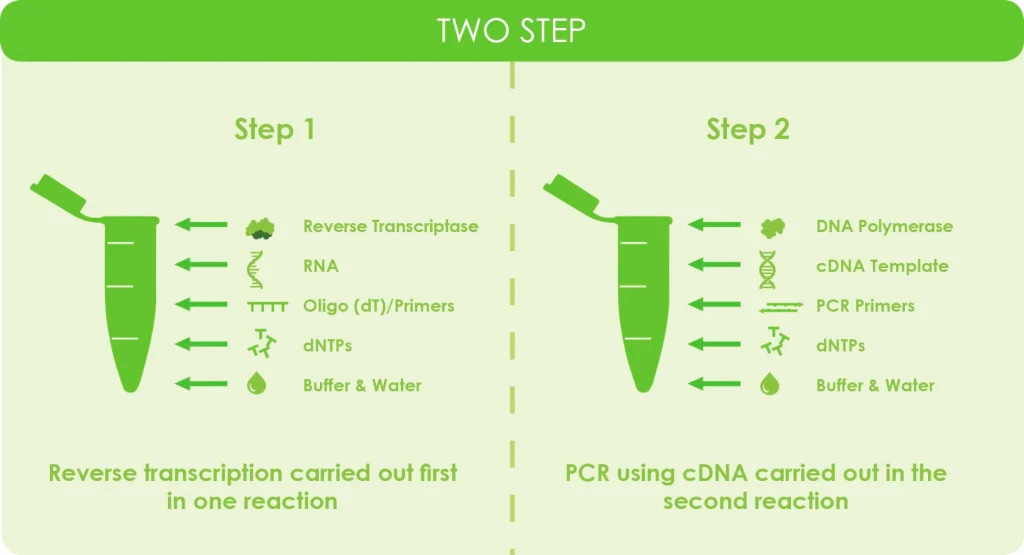
Για να κατανοήσουμε καλύτερα αυτή τη θεωρητική μετατροπή, ας ρίξουμε μια ματιά στα βήματα που εμπλέκονται στη σύνθεση του cDNA και ας δούμε τι μπορούμε να ανακαλύψουμε. Παρακάτω παρουσιάζονται τα 5 βήματα που παρέχει η Thermo Fisher για τη βέλτιστη δημιουργία cDNA. Εδώ θα μπορέσετε να δείτε τις πολυάριθμες μεταβολές που πρέπει να υποστεί το RNA καθώς και τις χημικές ουσίες που προστίθενται στο δείγμα για τη δημιουργία της επιθυμητής αντίδρασης:
5 Βήματα για τη Βέλτιστη Σύνθεση cDNA
"Η σύνθεση του DNA από ένα πρότυπο RNA, μέσω της αντίστροφης μεταγραφής, οδηγεί σε συμπληρωματικό DNA (cDNA). Το cDNA μπορεί στη συνέχεια να χρησιμεύσει ως πρότυπο σε μια ποικιλία μεταγενέστερων εφαρμογών για μελέτες RNA, όπως η γονιδιακή έκφραση- επομένως, η σύνθεση cDNA είναι το πρώτο βήμα για πολλά πρωτόκολλα στη μοριακή βιολογία. Εάν είστε νέος στη σύνθεση cDNA ή έμπειρος ερευνητής που επιθυμεί να βελτιστοποιήσει το πρωτόκολλό του, λάβετε υπόψη σας αυτά τα πέντε κρίσιμα βήματα που θα σας βοηθήσουν να διασφαλίσετε ότι η σύνθεση cDNA θα έχει τα υψηλότερα δυνατά αποτελέσματα.
Βήμα 1. Προετοιμασία δείγματος
Το RNA χρησιμεύει ως πρότυπο στη σύνθεση cDNA. Το ολικό RNA χρησιμοποιείται συνήθως στη σύνθεση cDNA για μεταγενέστερες εφαρμογές, όπως η RT-(q)PCR, ενώ συγκεκριμένοι τύποι RNA (π.χ. αγγελιοφόρο RNA (mRNA) και μικρά RNA, όπως το miRNA) μπορεί να εμπλουτιστούν για ορισμένες εφαρμογές, όπως η κατασκευή βιβλιοθήκης cDNA και η σκιαγράφηση προφίλ miRNA.
Η διατήρηση της ακεραιότητας του RNA είναι κρίσιμη και απαιτεί ειδικές προφυλάξεις κατά την εξαγωγή, την επεξεργασία, την αποθήκευση και την πειραματική χρήση. Οι βέλτιστες πρακτικές για την αποφυγή της υποβάθμισης του RNA περιλαμβάνουν τη χρήση γαντιών, τη διοχέτευση με σιφώνια με ρύγχη με φραγμό αερολύματος, τη χρήση εργαστηριακών σκευών και αντιδραστηρίων χωρίς νουκλεάση και την απολύμανση των χώρων εργασίας.
Για την απομόνωση και τον καθαρισμό του RNA, διατίθενται διάφορες στρατηγικές ανάλογα με τον τύπο των υλικών προέλευσης (π.χ. αίμα, ιστοί, κύτταρα, φυτά) και τους στόχους των πειραμάτων. Οι κύριοι στόχοι των ροών εργασίας απομόνωσης είναι η σταθεροποίηση των μορίων RNA, η αναστολή των RNασών και η μεγιστοποίηση της απόδοσης με κατάλληλες μεθόδους αποθήκευσης και εκχύλισης. Οι βέλτιστες μέθοδοι καθαρισμού απομακρύνουν ενδογενείς ενώσεις, όπως οι σύνθετοι πολυσακχαρίτες και το χουμικό οξύ από φυτικούς ιστούς που παρεμβαίνουν στη δραστηριότητα του ενζύμου- και τους κοινούς αναστολείς των αντίστροφων μεταγραφασών, όπως τα άλατα, τα ιόντα μετάλλων, η αιθανόλη και η φαινόλη. Μετά τον καθαρισμό, το RNA πρέπει να αποθηκεύεται στους -80°C με ελάχιστους κύκλους ψύξης-απόψυξης.
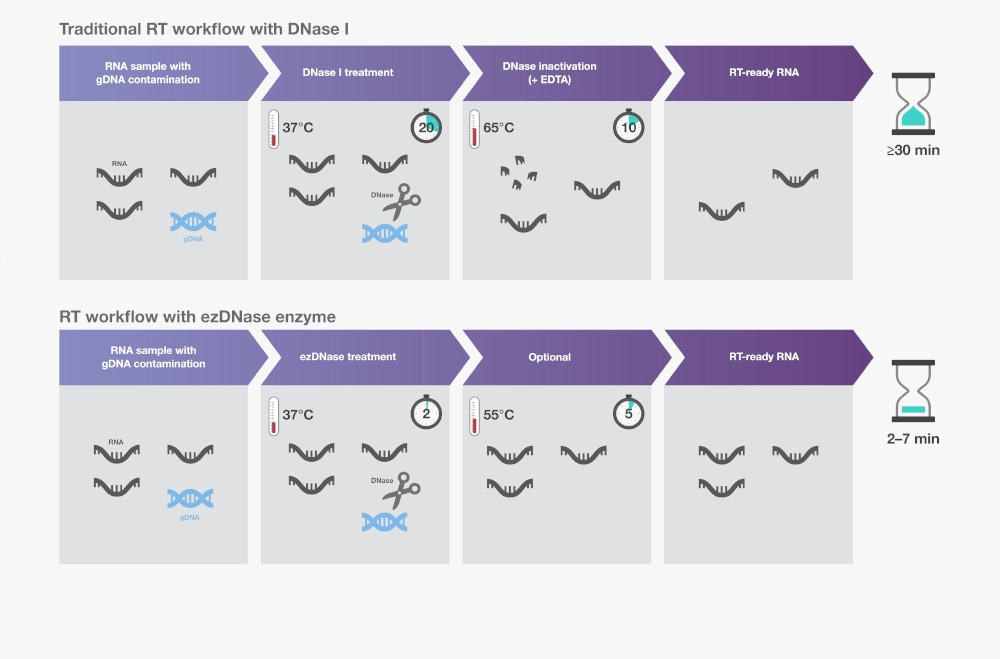
Βήμα 2. Απομάκρυνση του γονιδιωματικού DNA
Ίχνη γονιδιωματικού DNA (gDNA) μπορεί να καθαριστούν μαζί με το RNA. Το μολυσματικό gDNA μπορεί να επηρεάσει την αντίστροφη μεταγραφή και μπορεί να οδηγήσει σε ψευδώς θετικά αποτελέσματα, υψηλότερο υπόβαθρο ή χαμηλότερη ανίχνευση σε ευαίσθητες εφαρμογές όπως η RT-qPCR.
Η παραδοσιακή μέθοδος απομάκρυνσης του gDNA είναι η προσθήκη DNase I σε παρασκευάσματα απομονωμένου RNA. Η DNase I πρέπει να απομακρυνθεί πριν από τη σύνθεση cDNA, δεδομένου ότι τυχόν υπολείμματα ενζύμου θα αποικοδομούσαν το μονόκλωνο DNA. Δυστυχώς, μπορεί να προκύψει απώλεια ή βλάβη του RNA κατά τη διάρκεια της επεξεργασίας αδρανοποίησης με DNase I.
Ως εναλλακτική λύση στη DNάση Ι, διατίθενται ειδικές DNάσες διπλής αλυσίδας για την εξάλειψη του μολυσματικού gDNA χωρίς να επηρεάζεται το RNA ή τα μονόκλωνα DNA. Η θερμοδιαλυτή τους ιδιότητα επιτρέπει την απλή αδρανοποίηση σε σχετικά ήπια θερμοκρασία (π.χ. 55°C) χωρίς αρνητικές επιπτώσεις. Τέτοιες ειδικές για διπλές αλυσίδες, θερμοαπορροφήσιμες DNάσες μπορούν να επωαστούν με RNA για 2 λεπτά στους 37°C πριν από τις αντιδράσεις αντίστροφης μεταγραφής για τον εξορθολογισμό της ροής εργασίας (Εικόνα 1 ).
Βήμα 3. Επιλογή αντίστροφης μεταγραφάσης
Οι περισσότερες αντίστροφες μεταγραφάσες που χρησιμοποιούνται στη μοριακή βιολογία προέρχονται από το γονίδιο pol του ιού της μυελοβλάστης των πτηνών (AMV) ή του ιού της λευχαιμίας των ποντικών Moloney (MMLV). Η αντίστροφη μεταγραφάση του AMV ήταν ένα από τα πρώτα ένζυμα που απομονώθηκαν για τη σύνθεση cDNA στο εργαστήριο. Το ένζυμο διαθέτει ισχυρή δραστικότητα RNάσης Η που αποικοδομεί το RNA στα υβρίδια RNA:cDNA, με αποτέλεσμα την παραγωγή μικρότερων θραυσμάτων cDNA (<5 kb).
Η αντίστροφη μεταγραφάση MMLV έγινε μια δημοφιλής εναλλακτική λύση λόγω της μονομερούς δομής της, η οποία επέτρεπε απλούστερη κλωνοποίηση και τροποποιήσεις στο ανασυνδυασμένο ένζυμο. Παρόλο που η MMLV είναι λιγότερο θερμοσταθερή από την αντίστροφη μεταγραφάση AMV, η αντίστροφη μεταγραφάση MMLV είναι ικανή να συνθέτει μακρύτερο cDNA (<7 kb) με υψηλότερη απόδοση, λόγω της χαμηλότερης δραστικότητας της RNάσης H.
Για την περαιτέρω βελτίωση της σύνθεσης cDNA, η αντίστροφη μεταγραφάση MMLV έχει τροποποιηθεί για ακόμη χαμηλότερη δραστικότητα RNase H (δηλ. μεταλλαγμένη περιοχή RNase H ή RNaseH-), υψηλότερη θερμοσταθερότητα (έως 55°C) και αυξημένη επεξεργαστικότητα (65 φορές υψηλότερη). Αυτά τα χαρακτηριστικά έχουν ως αποτέλεσμα αυξημένο μήκος και απόδοση cDNA, υψηλότερη ευαισθησία, βελτιωμένη αντοχή στους αναστολείς και ταχύτερους χρόνους αντίδρασης (Πίνακας 1).
Βήμα 4. Προετοιμασία μίγματος αντίδρασης
Εκτός από το ένζυμο και τους εκκινητές, τα κύρια συστατικά της αντίδρασης για την αντίστροφη μεταγραφή περιλαμβάνουν το πρότυπο RNA (προεπεξεργασμένο για την απομάκρυνση του γονιδιωματικού DNA), ρυθμιστικό διάλυμα, dNTPs, DTT, αναστολέα RNάσης και νερό χωρίς RNάση (Εικόνα 2).

Βήμα 5. Εκτέλεση της σύνθεσης cDNA
Οι αντιδράσεις αντίστροφης μεταγραφής περιλαμβάνουν τρία βασικά βήματα: ανόπτηση εκκινητή, πολυμερισμό DNA και απενεργοποίηση ενζύμου. Η θερμοκρασία και η διάρκεια αυτών των βημάτων ποικίλλουν ανάλογα με την επιλογή εκκινητή, το RNA-στόχο και τη χρησιμοποιούμενη αντίστροφη μεταγραφάση.
Το κρίσιμο βήμα είναι κατά τη διάρκεια του πολυμερισμού του DNA. Σε αυτό το βήμα, η θερμοκρασία και η διάρκεια της αντίδρασης μπορεί να ποικίλλουν ανάλογα με την επιλογή του εκκινητή και τη χρησιμοποιούμενη αντίστροφη μεταγραφάση. Εάν χρησιμοποιείτε τυχαίους εξαμερείς, τότε συνιστούμε την επώαση της αντίδρασης αντίστροφης μεταγραφής σε θερμοκρασία δωματίου (~25 °C) για 10 λεπτά μετά την προσθήκη ενζύμου για την επέκταση των εκκινητών.
Μεταξύ των αντίστροφων μεταγραφασών υπάρχουν διαφορές στη θερμοσταθερότητα, η οποία με τη σειρά της καθορίζει την υψηλότερη βέλτιστη θερμοκρασία πολυμερισμού για την καθεμία. Η χρήση μιας θερμοσταθερής αντίστροφης μεταγραφάσης επιτρέπει, μια υψηλότερη θερμοκρασία αντίδρασης (π.χ. 50 °C), για να βοηθήσει στη μετουσίωση RNA με υψηλή περιεκτικότητα σε GC ή δευτερογενείς δομές χωρίς να επηρεαστεί η δραστικότητα του ενζύμου (Εικόνα 3). Με τέτοια ένζυμα, η επώαση σε υψηλή θερμοκρασία μπορεί να οδηγήσει σε αύξηση της απόδοσης, του μήκους και της αναπαράστασης του cDNA".

https://www.thermofisher.com/us/en/home/life-science/pcr/reverse-transcription/5steps-cDNA.html
Τώρα που έχουμε το γενικό περίγραμμα των βημάτων που απαιτούνται για τη σύνθεση cDNA από RNA, η Thermofisher τόνισε όμορφα ορισμένα από τα προβλήματα που μπορεί να προκύψουν κατά τη διαδικασία αυτή:
Ξεπερνώντας τα Εμπόδια στη Σύνθεση cDNA
"Μπορείτε να πείτε πολλά για ένα κύτταρο εξετάζοντας το RNA του, αλλά αυτά τα μεταβατικά μόρια είναι δύσκολο να μελετηθούν άμεσα. Το RNA αποικοδομείται εύκολα, επειδή η επιπλέον υδροξυλομάδα στο σάκχαρο ριβόζη είναι ιδιαίτερα αντιδραστική. Το DNA είναι πιο σταθερό, επειδή έχει κορμό σακχάρου δεοξυριβόζης και δίκλωνη δομή. Αυτός είναι ο λόγος για τον οποίο οι επιστήμονες χρησιμοποιούν την αντίστροφη μεταγραφή για να φτιάξουν συμπληρωματικό DNA (cDNA), το οποίο αποτυπώνει τις αλληλουχίες RNA σε μορφή DNA. Η παραγωγή cDNA είναι το πρώτο βήμα σε πολλές εφαρμογές μοριακής βιολογίας - από την κλωνοποίηση και την RT-PCR έως τις μικροσυστοιχίες και την αλληλούχιση επόμενης γενιάς.
Για να διατηρήσετε την εγκυρότητα των πειραματικών αποτελεσμάτων, χρειάζεστε cDNA που αναπαριστά πιστά το πρότυπο RNA. Τα σημαντικότερα εμπόδια στις αντιδράσεις σύνθεσης cDNA περιλαμβάνουν τις δευτερογενείς δομές RNA που μπορεί να επιβραδύνουν ή ακόμη και να σταματήσουν την αντίστροφη μεταγραφή. Η αποικοδόμηση του πρότυπου RNA από την εγγενή δραστηριότητα RNAάσης των αντίστροφων μεταγραφασών (RTs) είναι ένα άλλο πρόβλημα που οδηγεί σε περικομμένο cDNA. Επίσης, οι αναστολείς που υπάρχουν στα δείγματα RNA μπορεί να μειώσουν τη δραστηριότητα πολυμερισμού των ενζύμων της αντίστροφης μεταγραφάσης. Η αποικοδόμηση του προτύπου και ο αναποτελεσματικός πολυμερισμός έχουν ως αποτέλεσμα cDNA χαμηλής ποιότητας και ποσότητας.
Για να ενισχύσουν την απόδοση του ενζύμου, οι επιστήμονες της Thermo Fisher Scientific χρησιμοποίησαν τη μοριακή εξέλιξη και τον ορθολογικό σχεδιασμό για να κατασκευάσουν μια καλύτερη αντίστροφη μεταγραφάση. Ξεκινώντας με το γονίδιο RT άγριου τύπου από τον ιό της λευχαιμίας των ποντικών Moloney (M-MuLV), οι επιστήμονές μας εισήγαγαν μια σειρά βελτιώσεων για να σας βοηθήσουν να επιτύχετε καλύτερες αποδόσεις cDNA, μακρύτερα προϊόντα και μεγαλύτερη αναπαράσταση του RNA εισόδου.
Ενζυμική ενίσχυση #1: Αυξημένη θερμοσταθερότητα
Το μονόκλωνο RNA σχηματίζει βρόχους φουρκέτας και άλλες δευτερογενείς δομές, οι οποίες μπορούν να παρεμποδίσουν τη σύνθεση cDNA. Η ικανότητα μιας αντίστροφης μεταγραφάσης να ανέχεται υψηλές θερμοκρασίες μπορεί να βοηθήσει να ξεπεραστεί αυτή η πρόκληση. Η M-MuLV RT άγριου τύπου λειτουργεί στους 37-42°C, ενώ το τροποποιημένο ένζυμό μας διατηρεί τη δραστηριότητά του μέχρι τους 50-55°C. Οι αυξημένες θερμοκρασίες αντίδρασης αποσταθεροποιούν τις δευτερογενείς δομές του RNA, επιτρέποντας στην RT να διαβάσει την αλληλουχία, αυξάνοντας έτσι την απόδοση του cDNA.
Ενζυμική ενίσχυση #2: Μειωμένη δραστικότητα RNase H
Το πρώτο σκέλος της σύνθεσης cDNA δημιουργεί υβριδικά μόρια DNA-RNA. Οι RT έχουν συχνά ενσωματωμένη δραστηριότητα RNase H - την ικανότητα να υδρολύουν το RNA πριν από την ολοκλήρωση της δεύτερης αλυσίδας cDNA. Ωστόσο, η υπερβολική δραστηριότητα RNάσης μπορεί να αποικοδομήσει πρόωρα το πρότυπο RNA, γεγονός που μπορεί να μειώσει την απόδοση και το μήκος των προϊόντων cDNA. Η δική μας RT περιλαμβάνει τροποποιήσεις που μεταβάλλουν ή μειώνουν σημαντικά τη δραστικότητα της RNάσης H με αποτέλεσμα την αύξηση της απόδοσης των προϊόντων σύνθεσης cDNA πλήρους μήκους.
Ενζυμική ώθηση #3: Υψηλότερη επεξεργαστικότητα
Ουσίες που δεσμεύονται στο RNA και παρεμβαίνουν στη σύνθεση cDNA μεταφέρονται συνήθως από πηγές δειγμάτων RNA. Η επεξεργαστικότητα είναι ένα βασικό χαρακτηριστικό του ενζύμου που αναφέρεται στον αριθμό των νουκλεοτιδίων που ενσωματώνονται κατά τη διάρκεια ενός μεμονωμένου γεγονότος σύνδεσης. Η αυξημένη επεξεργαστικότητα συσχετίζεται με στενότερη πρόσδεση υποστρώματος και μπορεί να βελτιώσει την ανθεκτικότητα σε αυτούς τους αναστολείς. Η υψηλή επεξεργαστικότητα επιτρέπει στους RT να συνθέτουν μακρύτερες αλυσίδες cDNA σε μικρότερο χρόνο αντίδρασης. Για τη βελτιωμένη RT μας, χρειάζονται μόλις 15-30 λεπτά για τη σύνθεση cDNA, ανάλογα με το αν έχει αφαιρεθεί ή όχι το γονιδιωματικό DNA. Αυτή η αποτελεσματικότητα επιφέρει υψηλότερη απόδοση και ανώτερη ευαισθησία με δείγματα RNA χαμηλής ποιότητας ή χαμηλής ποσότητας, ακόμη και RNA από μεμονωμένα κύτταρα".
https://www.thermofisher.com/us/en/home/brands/thermo-scientific/molecular-biology/molecular-biology-learning-center/molecular-biology-resource-library/spotlight-articles/cdna-synthesis-overcome-hurdles.html
Από την παραπάνω πηγή φαίνεται ότι υπάρχουν μερικά πιθανά προβλήματα στη διαδικασία σύνθεσης cDNA, όπως:
Δευτερογενείς δομές RNA που μπορούν να σταματήσουν ή να σταματήσουν τη διαδικασία RT
Αποικοδόμηση του πρότυπου RNA από την εγγενή RNάση
Μειωμένος πολυμερισμός λόγω αναστολέων
Υπολείμματα που μεταφέρονται από πηγές δειγματοληψίας και παρεμβαίνουν στη σύνθεση cDNA

Ακόμη και με τον υποτιθέμενο "καθαρισμό" του RNA στα αρχικά στάδια εκχύλισης για τη δημιουργία του γονιδιώματος, εξακολουθούν να υπάρχουν προβλήματα που σχετίζονται με τη μόλυνση σε όλα τα υπόλοιπα στάδια αυτής της διαδικασίας. Στην RT-PCR, η επιμόλυνση είναι ένα επίμονο πρόβλημα που οδηγεί σε ανακριβή δεδομένα, καθώς η PCR δεν μπορεί να διαφοροποιήσει μεταξύ του συνθετικού cDNA και της επιμόλυνσης. Το πρόβλημα αυτό μπορεί μόνο να μετριαστεί (δηλαδή να γίνει λιγότερο σοβαρό) και παραμένει χωρίς εύκολη λύση:
Αποφυγή της Μόλυνσης του DNA στην RT-PCR
"Μια συχνή αιτία ανησυχίας μεταξύ των ερευνητών που εκτελούν ποσοτική RT-PCR είναι τα ανακριβή δεδομένα που οφείλονται σε επιμόλυνση DNA σε παρασκευάσματα RNA. Αν και η μόλυνση από DNA ανιχνεύεται εύκολα με την εκτέλεση ενός ελέγχου "χωρίς RT", δεν υπάρχει εύκολη θεραπεία. Σε αυτό το τεχνικό δελτίο, παρουσιάζουμε δεδομένα που δείχνουν τα επίπεδα μόλυνσης DNA σε RNA που παράγεται με διαφορετικές διαδικασίες και προτείνουμε διάφορα προληπτικά μέτρα που μπορούν να εφαρμοστούν για να μειωθεί ο αντίκτυπος αυτού του επίμονου προβλήματος.
RT-PCR και Γονιδιωματική Επιμόλυνση
Η RT-PCR είναι μια ολοένα και πιο δημοφιλής μέθοδος για την ποσοτική ανάλυση της γονιδιακής έκφρασης. Με αυτή τη δημοτικότητα έρχεται και η αυξημένη επίγνωση ότι οι περισσότερες τεχνικές που χρησιμοποιούνται για την απομόνωση ολικού RNA αποδίδουν RNA με σημαντικές ποσότητες επιμόλυνσης από γονιδιωματικό DNA. Η PCR δεν μπορεί να διακρίνει μεταξύ των στόχων cDNA που συντίθενται με αντίστροφη μεταγραφή και της επιμόλυνσης από γονιδιωματικό DNA".
http://diyhpl.us/~bryan/irc/protocol-online/protocol-cache/tb_176.html
Ένα άλλο ζήτημα που αντιμετωπίζει η RT-PCR είναι ότι αποτελεί την κύρια πηγή μεροληψίας και τεχνουργημάτων κατά την κατασκευή βιβλιοθηκών. Ο αριθμός των κύκλων που χρησιμοποιούνται μπορεί να επηρεάσει σε μεγάλο βαθμό την ποσότητα της προκατειλημμένης εικόνας που δημιουργείται, καθώς όσο μεγαλύτερος είναι ο κύκλος, τόσο μεγαλύτερη είναι η προκατειλημμένη εικόνα. Θεωρείται απαραίτητο να αποφεύγεται η μεροληψία, αλλά αυτό σπάνια συμβαίνει. Αυτές οι μεροληψίες και τα τεχνουργήματα εισέρχονται στο τελικό γονιδίωμα οδηγώντας σε ανακριβή, αναξιόπιστα και μη επαναλήψιμα αποτελέσματα:
Μεροληψία στην Προετοιμασία Βιβλιοθήκης RNA-seq: Τρέχουσες προκλήσεις και λύσεις
3.6. Αντίστροφη μεταγραφή
"Επί του παρόντος, οι στρατηγικές της ανάλυσης του μεταγραφώματος εξακολουθούν να μετατρέπουν το RNA σε cDNA πριν από την αλληλούχιση. Ένα γνωστό χαρακτηριστικό των αντίστροφων μεταγραφασών είναι ότι τείνουν να παράγουν ψευδές cDNA δεύτερης αλυσίδας μέσω της εξαρτώμενης από το DNA DNA πολυμεράσης. Αυτό μπορεί να μην είναι σε θέση να διακρίνει το sense και antisense μεταγράφημα και να δημιουργήσει δυσκολίες για την ανάλυση των δεδομένων".
3.7. Ενίσχυση με PCR
"Η PCR είναι ένα βασικό εργαλείο που χρησιμοποιείται ευρέως στα εργαστήρια μοριακής βιολογίας. Ειδικότερα, ο συνδυασμός της PCR και της αλληλούχισης NGS προώθησε την εκρηκτική ανάπτυξη της απόκτησης αλληλουχιών RNA. Ωστόσο, η ενίσχυση PCR έχει αποδειχθεί ότι είναι η κύρια πηγή τεχνουργημάτων και μεροληψίας της σύνθεσης των βάσεων κατά τη διαδικασία κατασκευής βιβλιοθήκης, η οποία μπορεί να οδηγήσει σε παραπλανητικά ή ανακριβή συμπεράσματα κατά την ανάλυση δεδομένων. Ως εκ τούτου, είναι απαραίτητο να αποφεύγεται η μεροληψία της PCR και έχουν καταβληθεί μεγάλες προσπάθειες για την προσπάθεια ελέγχου και μετριασμού της μεροληψίας στο ρεύμα".
3.8. Οι πηγές των μεροληψιών ενίσχυσης PCR και οι μέθοδοι βελτίωσης
3.8.1. Εξαιρετικά πλούσια AT/GC
"Μελέτες έχουν δείξει ότι θραύσματα ουδέτερου GC μπορούν να ενισχυθούν περισσότερο από θραύσματα πλούσια σε GC ή πλούσια σε AT. Ως εκ τούτου, τα θραύσματα με υψηλή περιεκτικότητα σε AT ή πολύ υψηλή περιεκτικότητα σε GC έχουν συχνά ελάχιστη ή καθόλου ενίσχυση [47, 48]. Αυτά τα δυσμενή χαρακτηριστικά έχουν ως αποτέλεσμα δυσκολίες στην αλληλούχιση γονιδιωμάτων εξαιρετικά πλούσιων σε ΑΤ, όπως το ανθρώπινο παράσιτο της ελονοσίας [48], ή γονιδιωμάτων με υψηλή περιεκτικότητα σε GC (Bordetella pertussis) (μέση περιεκτικότητα σε GC, περίπου 75%)".
3.8.2. Κύκλος PCR
"Όπως όλοι γνωρίζουμε, η PCR μπορεί να ενισχύσει εκθετικά τα πρότυπα DNA/cDNA, οδηγώντας έτσι σε σημαντική αύξηση της μεροληψίας ενίσχυσης με τον αριθμό των κύκλων PCR [51]. Ως εκ τούτου, συνιστάται η PCR να εκτελείται με τη χρήση όσο το δυνατόν λιγότερων αριθμών κύκλων για τον μετριασμό της μεροληψίας [52, 53]. Προς το παρόν, διάφορα εργαστήρια έχουν συγκρίνει διαφορετικούς αριθμούς κύκλων PCR για τη μείωση της μεροληψίας ενίσχυσης. Οι Wu et al. [54] πραγματοποίησαν μια ολοκληρωμένη ανάλυση. Τα αποτελέσματα της μελέτης υποδεικνύουν ότι σε σύγκριση με τον χαμηλότερο αριθμό κύκλων, ο υψηλότερος αριθμός κύκλων μπορεί να παράγει σημαντικές μεροληψίες ή τεχνουργήματα σε τυπικές ενισχύσεις μικτών προτύπων. Επιπλέον, η μελέτη των Sze και Schloss [55] έδειξε ότι η μείωση του αριθμού των κύκλων ενίσχυσης μπορεί επίσης να μειώσει τις μεροληψίες και τα τεχνουργήματα PCR χρησιμοποιώντας μια εικονική κοινότητα και δείγματα ανθρώπινων κοπράνων".
https://www.hindawi.com/journals/bmri/2021/6647597/
Τέλος, η ίδια η αντίστροφη μεταγραφάση αποτελεί πηγή μόλυνσης, γεγονός που δεν θα έπρεπε να είναι τόσο σοκαριστικό, αν αναλογιστεί κανείς το αρχικό υλικό που χρησιμοποιήθηκε για τη δημιουργία του ανασυνδυασμένου παρασκευάσματος. Στην εν λόγω μελέτη του 2011, διερευνήθηκαν πέντε από τα πιο συχνά χρησιμοποιούμενα ένζυμα RT και παρατηρήθηκε ότι όλα είχαν μη ειδική ενίσχυση cDNA. Ως αιτία του προβλήματος αυτού εικάζεται η εύρεση προσμίξεων RNA στην RT. Πρόκειται για ένα ζήτημα που περιγράφεται εκτενώς στην ιολογία και οδηγεί εύκολα σε ψευδώς θετικά αποτελέσματα, καθώς μπορεί να επηρεάσει την ειδικότητα της RT:
Η Εμπορική Αντίστροφη Μεταγραφάση ως πηγή ψευδώς θετικής ανίχνευσης RNA ειδικού για την αλυσίδα σε ανθρώπινα κύτταρα
"Μια συνήθως χρησιμοποιούμενη τεχνική για τη διερεύνηση της έκφρασης ενός αντι-συνδεδεμένου ncRNA είναι η ειδική για την αλυσίδα αντίστροφη μεταγραφή σε συνδυασμό με την αλυσιδωτή αντίδραση πολυμεράσης (RT-PCR). Το πλεονέκτημα αυτής της ακριβούς τεχνικής είναι ότι δεν απαιτεί ειδικό εξοπλισμό ή τεχνογνωσία. Το μειονέκτημα είναι ότι μπορεί εύκολα να οδηγήσει σε ψευδώς θετικά αποτελέσματα. Εφαρμόσαμε την ειδική για την αλυσίδα RT-PCR για να διερευνήσουμε την παρουσία αντισημαστικού ncRNA που σχετίζεται με τον υποδοχέα ρετινοϊκού οξέος βήτα 2 (RARβ2) σε διάφορες ανθρώπινες καρκινικές κυτταρικές σειρές. Με την εκτέλεση αυτής της τεχνικής, παρατηρήσαμε ψευδώς θετική ανίχνευση του ncRNA. Για την ακριβή ερμηνεία των αποτελεσμάτων σε πειράματα RT-PCR, εισαγάγαμε έναν έλεγχο "Χωρίς εκκινητή" που αποκαλύπτει τη μη ειδική σύνθεση cDNA. Επιπλέον, αναφέρουμε την παρουσία μη ειδικής ενίσχυσης cDNA με πέντε από τις πιο συχνά χρησιμοποιούμενες αντίστροφες μεταγραφάσες απουσία προστιθέμενων εκκινητών. Διαπιστώσαμε ότι η επιλογή της αντίστροφης μεταγραφάσης καθώς και οι συνθήκες της αντίδρασης (θερμοκρασία RT και αριθμός κύκλων PCR) είναι σημαντικές παράμετροι για την επιλογή, καθώς οι διαφορετικές αντίστροφες μεταγραφάσες δεν παρουσιάζουν το ίδιο υπόβαθρο σύνθεσης cDNA. Αυτό το φαινόμενο που είχε παρατηρηθεί προηγουμένως αναφέρθηκε ότι προέρχεται από την "αυτοπροστασία" του προτύπου RNA. Εδώ, αναφέρουμε μάλλον την παρουσία προσμίξεων RNA που σχετίζονται με μία από τις αντίστροφες μεταγραφάσες που μελετήθηκαν και οι οποίες ενδέχεται να συμβάλλουν στη μη ειδική σύνθεση cDNA".
"Το κρίσιμο βήμα της διαδικασίας είναι η παραγωγή DNA από το πρότυπο RNA από την αντίστροφη μεταγραφάση του ρετροϊού (RT) παρουσία ειδικών για την αλληλουχία ή τυχαίων ολιγοδεοξυνουκλεοτιδικών εκκινητών. Για την ειδική ταυτοποίηση αντι-αναληπτικών ncRNA που επικαλύπτουν μεταγράμματα αίσθησης αλλά είναι αντίθετα προσανατολισμένα, δεν είναι δυνατή η χρήση τυχαίων ολιγοδεοξυνουκλεοτιδικών εκκινητών RT-PCR, δεδομένου ότι δεν μπορεί να κάνει διάκριση μεταξύ των δύο ειδών. Για να αποφευχθεί αυτό το πρόβλημα, χρησιμοποιούνται εκκινητές ειδικοί για την αλυσίδα που οδηγούν στην ειδική αντίστροφη μεταγραφή του λογικού ή του αντι-νοηματικού RNA και στη συνέχεια στην ενίσχυση με PCR. Για την ανίχνευση του. αντισημικού RNA, προστίθεται στην αντίδραση RT μόνο ο εκκινητής που είναι συμπληρωματικός προς αυτή την αλυσίδα, ώστε να επιτευχθεί σύνθεση cDNA μόνο όταν το αντισημικό μεταγράφημα είναι παρόν στο συνολικό δείγμα RNA. Εδώ εφαρμόσαμε αυτή την τεχνική σε έναν επιλεγμένο στόχο και παρατηρήσαμε μη ειδική ενίσχυση και ψευδώς θετικά αποτελέσματα. Τα αποτελέσματα αυτά προέκυψαν με υψηλή αναπαραγωγιμότητα με πέντε διαφορετικές εμπορικές RT σε τρία γονίδια σε διάφορες ανθρώπινες κυτταρικές σειρές. Αυτή η μη ειδική σύνθεση cDNA που οδηγεί σε αλλοπρόσαλλη ανίχνευση με ειδικότητα στην αλυσίδα είχε παρατηρηθεί προηγουμένως στον τομέα της ιολογίας, όπου η προσέγγιση αυτή χρησιμοποιήθηκε για τη διάκριση μεταξύ αρνητικής και θετικής αλυσίδας ιικού RNA [13e16] και είναι γνωστή ως "false-priming". Διάφοροι μηχανισμοί έχουν προταθεί για να εξηγήσουν πώς η σύνθεση cDNA προέρχεται από false-priming κατά τη διάρκεια της αντίδρασης RT, συμπεριλαμβανομένου του τυχαίου priming από μολυσματικά ενδογενή ή εξωγενή νουκλεϊκά οξέα [14,17,18] και των δευτερογενών δομών φουρκέτας του RNA που μπορούν να αναγνωριστούν και να επεκταθούν από τα ένζυμα RT, το λεγόμενο "self-priming" [17,19,20]. Εδώ αποδεικνύουμε ότι αυτή η μη ειδική σύνθεση cDNA είναι ένα παγκόσμιο φαινόμενο που συμβαίνει και με το ανθρώπινο κυτταρικό RNA. Ανάλογα με το χρησιμοποιούμενο ένζυμο RT, οδηγεί σε διαφορετικά επίπεδα ψευδώς θετικών αποτελεσμάτων RT-PCR για συγκεκριμένη αλυσίδα, τα οποία μπορούν να παρερμηνευθούν ελλείψει κατάλληλων ελέγχων. Αυτό είναι θεμελιώδους σημασίας σε όλες τις εφαρμογές που αναζητούν ανίχνευση ειδικής για την αλυσίδα sense και antisense RNA. Στα πειράματά μας, βρήκαμε μικρούς προσμίξεις RNA σε εμπορικά παρασκευάσματα AMV RT που θα μπορούσαν να εμπλέκονται στη σύνθεση cDNA με λανθασμένη προετοιμασία".
"Κατά τη διάρκεια της έρευνάς μας σχετικά με την ύπαρξη των αντι-αντι-νοηματικών ncRNAs RARb2, συναντήσαμε ένα απροσδόκητα υψηλό επίπεδο ψευδώς θετικών αποτελεσμάτων. Η σύνθεση cDNA από κυτταρικό ολικό RNA χωρίς την προσθήκη εκκινητών αποκάλυψε μια συνολική μη ειδική αντίστροφη μεταγραφή του προτύπου RNA. Αυτό το γεγονός ψευδούς εκκίνησης έχει τεκμηριωθεί επαρκώς στον τομέα της ιολογίας, στον οποίο αποτελεί υψηλή πηγή ψευδώς θετικών αποτελεσμάτων κατά την ανίχνευση ιικού RNA ειδικής αλυσίδας [13e16]. Πιστεύεται ότι η προέλευση της ανεξάρτητης από εκκινητή σύνθεσης cDNA προέρχεται είτε από την παρουσία κυτταρικών επιμολυντών νουκλεϊκών οξέων, όπως το oligo(dT) [18], είτε από εξωγενείς επιμολυντές νουκλεϊκών οξέων [14,17] είτε από δευτερογενώς δομημένα RNA [17,19,20]".
"Αξίζει να σημειωθεί ότι δείξαμε ότι πέντε από τα πιο συχνά χρησιμοποιούμενα ένζυμα RT εμφανίζουν μη ειδική σύνθεση cDNA, αλλά η ποσότητα του υποβάθρου δεν ήταν η ίδια ανάλογα με το ένζυμο RT (Εικ. 3). Ως εκ τούτου, η παρατήρηση αυτή έθεσε το ενδεχόμενο η μη ειδική σύνθεση cDNA να προέρχεται από το ίδιο το ένζυμο και όχι από το πρότυπο RNA. Αποδείξαμε ότι δεν εμπλέκεται η δραστηριότητα της RNάσης H της αντίστροφης μεταγραφάσης, δεδομένου ότι η σύνθεση cDNA παρατηρήθηκε ανεξάρτητα από μια σχετική δραστηριότητα RNάσης H (Εικ. 3)".
"Όπως περιγράφηκε εκτενώς στον ιό, δείξαμε ότι η έναρξη της σύνθεσης cDNA κατά την RT-PCR μπορεί να συμβεί σε ανθρώπινο κυτταρικό mRNA χωρίς την προσθήκη εξωγενών εκκινητών στο στάδιο της RT. Αυτή η μη ειδική σύνθεση cDNA είναι ένα παγκόσμιο φαινόμενο, το οποίο ανιχνεύεται σε διαφορετικό επίπεδο με πέντε από τις πλέον χρησιμοποιούμενες εμπορικές αντίστροφες τρανσκριπτάσες. Αυτό μπορεί να παρεμβαίνει στην ειδικότητα της RT και αποτελεί υψηλή πηγή ψευδώς θετικών αποτελεσμάτων, ιδίως κατά τη διάκριση του αντισημειακού από το λογικό RNA".
https://pubmed.ncbi.nlm.nih.gov/21689721/

Συνοπτικά:
Η αντίληψη ότι η πληροφορία μπορεί πάντα να ρέει από το DNA στο RNA στην πρωτεΐνη αναφέρθηκε κάπως χαριτολογώντας ως το Κεντρικό Δόγμα της μοριακής βιολογίας
Ωστόσο, αυτό το κεντρικό δόγμα προκάλεσε τους επιστήμονες να βρουν εξαιρέσεις σε αυτόν τον κανόνα
Οι ιολόγοι βρήκαν την εξαίρεση όταν ανακαλύφθηκε ότι οι "ρετροϊοί" διέθεταν μηχανισμούς "αντίστροφης μεταγραφής".
Αυτό σημαίνει ότι μπορούν να παίρνουν αλυσίδες RNA και να παράγουν αλυσίδες DNA από αυτές και έτσι η πληροφορία ρέει αντίστροφα από το RNA πίσω στο DNA (πόσο βολικό...)
Το DNA που προκύπτει από αυτή τη διαδικασία ονομάζεται συμπληρωματικό DNA (cDNA)
Το cDNA παράγεται είτε από κάποιους "ιούς" είτε συντίθεται στα εργαστήρια
Το cDNA μπορεί να περιγραφεί ως gDNA χωρίς όλες τις απαραίτητες μη κωδικοποιητικές περιοχές
Οι επιστήμονες χρησιμοποιούν "ιϊκά" ένζυμα για να παρασκευάσουν cDNA από RNA που απομονώθηκε από τα κύτταρα και τους ιστούς που μελετούν
Για να απομονωθεί cDNA, πρέπει πρώτα να απομονωθεί το RNA ενός οργανισμού (κάτι που δεν γίνεται ποτέ με τους "ιούς")
Στη συνέχεια, χρησιμοποιώντας ένα ένζυμο αντίστροφης μεταγραφάσης (ένα ένζυμο που υποτίθεται ότι απομονώθηκε από έναν "ιό" που δεν απομονώθηκε ποτέ) μπορεί να παραχθεί cDNA
Αυτή είναι η διαδικασία που χρησιμοποιούν οι "ρετροϊοί" για να ενσωματωθούν στα κύτταρα του ξενιστή τους (με άλλα λόγια, η ύπαρξη του cDNA βασίζεται σε μη παρατηρημένες υποθετικές διαδικασίες ανύπαρκτων "ιών").
Αυτό είναι δυνατό επειδή οι "ρετροϊοί" χρησιμοποιούν RNA ως γονιδιωματικό τους υλικό αντί για DNA, και αυτό μεταγράφεται αντίστροφα στο cDNA, το οποίο στη συνέχεια υφίσταται κανονική μεταγραφή και οδηγεί στην "ιική" πρωτεΐνη στον ξενιστή
Για παράδειγμα, η αλληλουχία του γονιδιώματος του "SARS-COV-2", καθώς και οι άκρες του, προσδιορίστηκαν και επιβεβαιώθηκαν με PCR αντίστροφης μεταγραφής (RT-PCR)10 και ταχεία ενίσχυση των άκρων του cDNA 5′/3′ (RACE).
Στη συνέχεια κατασκευάστηκε βιβλιοθήκη RNA με τη χρήση του κιτ SMARTer Stranded Total RNA-Seq v.2 (TaKaRa)
Χρησιμοποιήθηκε η SMARTScribe RT, η οποία είναι μια τροποποιημένη αντίστροφη μεταγραφάση Moloney Murine Leukemia "Virus"
Η παντεταριασμένη διαδικασία καθαρισμού και τα αυστηρά πρότυπα ποιοτικού ελέγχου διασφαλίζουν ότι έχουν απομακρυνθεί σχεδόν όλες (δηλαδή όχι όλες) οι μολυσματικές νουκλεάσες
Με άλλα λόγια, η RT που χρησιμοποιήθηκε για τη μετατροπή του RNA "SARS-COV-2" σε cDNA δημιουργήθηκε συνθετικά σε εργαστήριο με "απομόνωση" από E. coli που εκφράζει ένα τμήμα του γονιδίου pol του "ιού" της λευχαιμίας του ποντικού σε ένα πλασμιδιακό γονίδιο
Αν και έχει παραχθεί ανασυνδυασμένος MMLV-RT σε Escherichia coli (Tanese et al. 1985- Roth et al. 1985- Kotewicz et al. 1985), οι συνθήκες έκφρασης και καθαρισμού δεν αναλύθηκαν λεπτομερώς.
Μια βιβλιοθήκη αλληλούχισης μπορεί να δημιουργηθεί ξεκινώντας από γονιδιωματικό DNA ή από RNA
Σε κάθε βιβλιοθήκη αλληλούχισης RNA υπάρχει ένα επιπλέον βήμα: η μετατροπή του RNA σε cDNA
Η PCR αντίστροφης μεταγραφής επιτρέπει τη χρήση του RNA ως προτύπου για τη δημιουργία συμπληρωματικού DNA (cDNA)
Το cDNA μπορεί στη συνέχεια να χρησιμεύσει ως πρότυπο σε διάφορες μεταγενέστερες εφαρμογές για μελέτες RNA, όπως η γονιδιακή έκφραση- ως εκ τούτου, η σύνθεση cDNA αποτελεί το πρώτο βήμα για πολλά πρωτόκολλα στη μοριακή βιολογία
5 βήματα για τη βέλτιστη σύνθεση cDNA
Προετοιμασία δείγματος
○ Η διατήρηση της ακεραιότητας του RNA είναι κρίσιμη και απαιτεί ειδικές προφυλάξεις κατά την εξαγωγή, την επεξεργασία, την αποθήκευση και την πειραματική χρήση
○ Οι κύριοι στόχοι των ροών εργασίας απομόνωσης είναι η σταθεροποίηση των μορίων RNA, η αναστολή των RNασών και η μεγιστοποίηση της απόδοσης με κατάλληλες μεθόδους αποθήκευσης και εκχύλισης
Απομάκρυνση του γονιδιωματικού DNA
○ Ίχνη γονιδιωματικού DNA (gDNA) μπορούν να καθαριστούν μαζί με το RNA
○ Το μολυσματικό gDNA μπορεί να επηρεάσει την αντίστροφη μεταγραφή και μπορεί να οδηγήσει σε ψευδώς θετικά αποτελέσματα, υψηλότερο υπόβαθρο ή χαμηλότερη ανίχνευση σε ευαίσθητες εφαρμογές όπως η RT-qPCR
○ Η παραδοσιακή μέθοδος απομάκρυνσης του gDNA είναι η προσθήκη DNase I σε παρασκευάσματα απομονωμένου RNA
○ Δυστυχώς, μπορεί να προκύψει απώλεια ή βλάβη του RNA κατά τη διάρκεια της επεξεργασίας αδρανοποίησης με DNase I
Επιλογή αντίστροφης μεταγραφάσης
○ Οι περισσότερες αντίστροφες μεταγραφάσες που χρησιμοποιούνται στη μοριακή βιολογία προέρχονται από το γονίδιο pol του "ιού της μυελοβλάστης των πτηνών" (AMV) ή του "ιού της λευχαιμίας των ποντικών Moloney" (MMLV)
Προετοιµασία µείγµατος αντίδρασης
○ Εκτός από το ένζυμο και τους εκκινητές, τα κύρια συστατικά της αντίδρασης για την αντίστροφη μεταγραφή περιλαμβάνουν το πρότυπο RNA (προεπεξεργασμένο για την απομάκρυνση του γονιδιωματικού DNA), ρυθμιστικό διάλυμα, dNTPs, DTT, αναστολέα RNάσης και νερό χωρίς RNάση.
Σύνθεση cDNA
○ Οι αντιδράσεις αντίστροφης μεταγραφής περιλαμβάνουν τρία κύρια βήματα: ανόπτηση εκκινητή, πολυμερισμό DNA και απενεργοποίηση ενζύμου
Το RNA αποικοδομείται εύκολα επειδή η επιπλέον υδροξυλομάδα στο σάκχαρο ριβόζης είναι ιδιαίτερα αντιδραστική
Για να διατηρήσετε την εγκυρότητα των πειραματικών αποτελεσμάτων, χρειάζεστε cDNA που να αντιπροσωπεύει πιστά το πρότυπο RNA
Τα σημαντικότερα εμπόδια στις αντιδράσεις σύνθεσης cDNA περιλαμβάνουν:
Οι δευτερογενείς δομές του RNA που μπορεί να επιβραδύνουν ή ακόμη και να σταματήσουν την αντίστροφη μεταγραφή
Η αποικοδόμηση του πρότυπου RNA από την εγγενή δραστηριότητα RNAάσης των αντίστροφων μεταγραφασών (RT)
Οι αναστολείς που υπάρχουν στα δείγματα RNA μπορεί να μειώσουν τη δραστηριότητα πολυμερισμού των ενζύμων της αντίστροφης μεταγραφάσης
Η αποικοδόμηση του προτύπου και ο αναποτελεσματικός πολυμερισμός έχουν ως αποτέλεσμα cDNA χαμηλής ποιότητας και ποσότητας
Το μονόκλωνο RNA σχηματίζει βρόχους φουρκέτας και άλλες δευτερογενείς δομές, οι οποίες μπορούν να παρεμποδίσουν τη σύνθεση cDNA
Η υπερβολική δραστηριότητα RNάσης μπορεί να αποικοδομήσει πρόωρα το πρότυπο RNA, γεγονός που μπορεί να μειώσει την απόδοση και το μήκος των προϊόντων cDNA
Ουσίες που δεσμεύονται στο RNA και παρεμβαίνουν στη σύνθεση cDNA μεταφέρονται συνήθως από πηγές δειγμάτων RNA
Μια συχνή αιτία ανησυχίας μεταξύ των ερευνητών που εκτελούν ποσοτική RT-PCR είναι τα ανακριβή δεδομένα που οφείλονται σε επιμόλυνση DNA στα παρασκευάσματα RNA.
Δεν υπάρχει εύκολη λύση και οι διαδικασίες μπορούν μόνο να μειώσουν τον αντίκτυπο αυτού του επίμονου προβλήματος
Οι περισσότερες τεχνικές που χρησιμοποιούνται για την απομόνωση ολικού RNA αποδίδουν RNA με σημαντικές ποσότητες μόλυνσης από γονιδιωματικό DNA
Η PCR δεν μπορεί να διακρίνει μεταξύ των στόχων cDNA που συντίθενται με αντίστροφη μεταγραφή και της μόλυνσης από γονιδιωματικό DNA
Ένα γνωστό χαρακτηριστικό των αντίστροφων μεταγραφασών είναι ότι τείνουν να παράγουν ψευδές cDNA δεύτερης αλυσίδας μέσω της DNA-εξαρτώμενης DNA πολυμεράσης
Η ενίσχυση με PCR έχει αποδειχθεί ότι είναι η κύρια πηγή τεχνουργημάτων και μεροληψίας της σύνθεσης των βάσεων κατά τη διαδικασία κατασκευής βιβλιοθήκης, η οποία μπορεί να οδηγήσει σε παραπλανητικά ή ανακριβή συμπεράσματα κατά την ανάλυση των δεδομένων
Πηγές μεροληψίας ενίσχυσης PCR:
Εξαιρετικά πλούσια σε AT/GC
○ Μελέτες έχουν δείξει ότι θραύσματα ουδέτερου GC μπορούν να ενισχυθούν περισσότερο από θραύσματα πλούσια σε GC ή πλούσια σε AT
○ Επομένως, τα θραύσματα με υψηλή περιεκτικότητα σε AT ή πολύ υψηλή περιεκτικότητα σε GC συχνά έχουν ελάχιστη ή καθόλου ενίσχυση
Κύκλοι PCR
○ Ο υψηλότερος αριθμός κύκλων μπορεί να προκαλέσει σημαντικές μεροληψίες ή τεχνουργήματα στις τυπικές ενισχύσεις μικτών προτύπων
○ Η μείωση του αριθμού των κύκλων της ενίσχυσης μπορεί να μειώσει (όχι να εξαλείψει) τις μεροληψίες και τα τεχνουργήματα της PCR
Το μειονέκτημα της RT-PCR είναι ότι μπορεί να οδηγήσει εύκολα σε ψευδώς θετικά αποτελέσματα
Μια μελέτη του 2011 ανέφερε με υψηλή αναπαραγωγιμότητα την παρουσία μη ειδικής ενίσχυσης cDNA με πέντε από τις πιο συχνά χρησιμοποιούμενες αντίστροφες μεταγραφάσες σε τρία γονίδια σε διάφορες ανθρώπινες κυτταρικές σειρές χωρίς την προσθήκη εκκινητών.
Ανέφεραν ότι η παρουσία προσμίξεων RNA που σχετίζονται με μία από τις αντίστροφες μεταγραφάσες που μελετήθηκαν μπορεί να συμβάλει στη μη ειδική σύνθεση cDNA
Αυτή η μη ειδική σύνθεση cDNA που οδηγεί σε αλλοπρόσαλλη ανίχνευση με ειδικότητα στην αλυσίδα είχε παρατηρηθεί προηγουμένως στον τομέα της ιολογίας, όπου η προσέγγιση αυτή χρησιμοποιήθηκε για τη διάκριση μεταξύ αρνητικής και θετικής αλυσίδας "ιικού" RNA και είναι γνωστή ως "ψευδής εκκίνηση" (false-priming)
Διάφοροι μηχανισμοί έχουν προταθεί για να εξηγήσουν πώς η σύνθεση cDNA προέρχεται από ψευδο-πρωτογενή εκχύλιση κατά τη διάρκεια της αντίδρασης RT, συμπεριλαμβανομένης της τυχαίας εκχύλισης από μολυσματικά ενδογενή ή εξωγενή νουκλεϊκά οξέα και δευτερογενείς δομές φουρκέτας RNA που μπορούν να αναγνωριστούν και να επεκταθούν από τα ένζυμα RT, η λεγόμενη "αυτο-εκκίνηση"(«self-priming»)
Αυτό το φαινόμενο ψευδούς εκκίνησης έχει τεκμηριωθεί καλά στον τομέα της ιολογίας, στον οποίο αποτελεί υψηλή πηγή ψευδώς θετικών αποτελεσμάτων κατά την ανίχνευση "ιικού" RNA ειδικής αλυσίδας
Πιστεύεται ότι η προέλευση της σύνθεσης cDNA χωρίς εκκινητή προέρχεται από:
Την παρουσία κυτταρικών νουκλεϊκών οξέων προσμίξεων όπως το oligo(dT),
Εξωγενείς (από έξω από εμάς) προσμίξεις νουκλεϊκών οξέων
Δευτερογενώς δομημένα RNA
Η παρατήρηση ότι η ποσότητα του υποβάθρου δεν ήταν η ίδια ανάλογα με το ένζυμο RT έθεσε το ενδεχόμενο η μη ειδική σύνθεση cDNA να προέρχεται από το ίδιο το ένζυμο και όχι από το πρότυπο RNA.
Όπως περιγράφεται εκτενώς στο "ιός", έδειξαν ότι η έναρξη της σύνθεσης cDNA κατά τη διάρκεια της RT-PCR μπορεί να συμβεί σε ανθρώπινο κυτταρικό mRNA χωρίς την προσθήκη εξωγενών εκκινητών στο στάδιο της RT
Αυτό μπορεί να επηρεάσει την εξειδίκευση της RT και αποτελεί υψηλή πηγή ψευδώς θετικών αποτελεσμάτων, ιδίως στη διάκριση του αντισημειακού από το λογικό RNA

Είναι εύκολα αντιληπτό ότι η μετατροπή του RNA σε cDNA προκειμένου να δημιουργηθεί μια βιβλιοθήκη αλληλούχισης είναι εξίσου γεμάτη με ζητήματα που αφορούν τη μόλυνση, τις μεροληψίες, τη δημιουργία τεχνουργημάτων, τα σφάλματα αλληλούχισης, τα ψευδή αποτελέσματα κ.λπ. όπως και τα προηγούμενα βήματα που σχετίζονται με τον "καθαρισμό" και τον κατακερματισμό του RNA. Η όλη διαδικασία στηρίζεται σε ένα ένζυμο της αντίστροφης μεταγραφάσης που ελήφθη συνθετικά από το γονίδιο pol είτε του σαρκώματος των πτηνών είτε του "ιού" της λευχαιμίας του ποντικού. Προφανώς, για να είναι αυτό ακριβές, απαιτείται πρώτα ο καθαρισμός και η απομόνωση ενός από τους δύο αυτούς "ιούς" προκειμένου να απομονωθεί ένα συγκεκριμένο γονίδιο ενός ενζύμου που υποτίθεται ότι δημιουργήθηκε από αυτούς. Καθώς κανένας "ιός" δεν έχει ποτέ καθαριστεί και απομονωθεί σωστά, το ένζυμο RT που είναι κρίσιμο για τη δημιουργία του cDNA που απαιτείται για την αλληλουχία ενός "ιού" είναι εξίσου υποθετικό με τους "ιούς" που αλληλουχίζονται.
Είναι γνωστό ότι η μέθοδος RT-PCR είναι επιρρεπής σε ψευδή αποτελέσματα, μεροληψίες, τεχνουργήματα και επιμόλυνση. Ωστόσο, το βήμα αυτό είναι ζωτικής σημασίας, καθώς υποτίθεται ότι δημιουργεί το υλικό cDNA που χρησιμοποιείται για την αλληλούχιση. Πρέπει να πιστέψουμε ότι τα πολυάριθμα βήματα που εμπλέκονται σε αυτή τη διαδικασία δημιουργίας γονιδιώματος παραμένουν εντελώς απαλλαγμένα από επιμολύνσεις/μεροληψίες/λάθη, ενώ αυτό προφανώς δεν ισχύει, όπως παραδέχονται διάφορες πηγές και μελέτες; Είναι αυτό έστω και μια ρεαλιστική προσδοκία και αποτέλεσμα γνωρίζοντας τα πολυάριθμα βήματα που εμπλέκονται και τους διάφορους τρόπους με τους οποίους τα πράγματα μπορούν να πάνε, και τακτικά πάνε, στραβά; Κάθε πρόσθετη διαδικασία στην οποία υποβάλλεται το αρχικό δείγμα, όπως η εξαγωγή του RNA, ο κατακερματισμός του RNA και η μετατροπή του RNA σε cDNA, είναι ένα ακόμη βήμα πιο μακριά από την πραγματικότητα. Κάθε αντίδραση είναι μη παρατηρήσιμη, εξηγείται υποθετικά και εξαρτάται εξ ολοκλήρου από την περιορισμένη τεχνολογία και την πεποίθηση ότι κάθε βήμα πραγματοποιείται κατάλληλα και με ακρίβεια χωρίς σφάλματα. Λαμβάνοντας υπόψη όλες τις πληροφορίες, το μόνο λογικό συμπέρασμα που μπορεί να εξαχθεί είναι ότι η μετατροπή του RNA σε cDNA μέσω της RT-PCR είναι εξίσου υποθετική με τους "ιούς" που λέγεται ότι έχουν αλληλουχία και τα ένζυμα που χρησιμοποιούνται για τη δημιουργία τους.


Αποχαιρετισμός στην Ιολογία - Έκθεση Eμπειρογνώμονα (Expert Edition)

Μετάφραση: Απολλόδωρος 15 Σεπτεμβρίου 2022 | Dr. Mark Bailey Διαβάστε το εδώThanks for reading Apollodoros’s Newsletter! Subscribe for free to receive new posts and support my work.
---Δικτυογραφία :
The Hypothetical Conversion of RNA to cDNA – ViroLIEgy
https://viroliegy.com/2022/02/19/the-hypothetical-conversion-of-rna-to-cdna/