


ΦΑΚΕΛΛΟΣ "ΙΟΛΟΓΙΑ": Διάτμηση/Kατακερματισμός DNA και RNA


Μετάφραση: Απολλόδωρος
15 Φεβρουαρίου 2022 | ViroLIEgy | Διαβάστε το εδώ.
Υπάρχουν πολυάριθμα βήματα που ακολουθούνται για τη δημιουργία ενός γονιδιώματος, είτε πρόκειται για ανθρώπινο, ζωικό, βακτηριακό ή "ιογενές". Συχνά είναι πολύπλοκα και απαιτούν διάφορες μεθόδους και τεχνολογίες, όπως ενίσχυση PCR και αλληλουχία, προκειμένου να προκύψει το τελικό προϊόν. Καθώς η διαδικασία είναι πολύ περίπλοκη και πολύπλοκη, υπάρχουν πολλαπλοί τρόποι με τους οποίους η ακρίβεια και η αξιοπιστία του γονιδιώματος μπορεί να τεθεί σε κίνδυνο στην πορεία. Αυτό μπορεί να παρατηρηθεί στην αρχή της διαδικασίας κατά το κρίσιμο βήμα της διάτμησης του DNA/RNA που λαμβάνεται από ιστούς ή κυτταρικές καλλιέργειες στο επιθυμητό εύρος θραυσμάτων. Αυτό γίνεται συνήθως είτε με φυσικά είτε με χημικά μέσα και αποτελεί απόλυτη απαίτηση μετά την εξαγωγή του RNA προκειμένου να δημιουργηθεί η βέλτιστη βιβλιοθήκη για το δείγμα που πρόκειται να αλληλουχηθεί. Το μέγεθος των θραυσμάτων εξαρτάται από την τεχνολογία που επιλέγεται και χρησιμοποιείται. Εάν προκύψουν προβλήματα κατά τον κατακερματισμό, αυτό μπορεί να εισάγει μεροληψίες, τεχνουργήματα και σφάλματα που θα πολλαπλασιαστούν και θα επιδεινωθούν στη συνέχεια στο τελικό γονιδίωμα.
Οι πληροφορίες που παρουσιάζονται παρακάτω έχουν συγκεντρωθεί από διάφορες πηγές και μπορεί να φαίνονται αρχικά συγκεχυμένες. Παρουσιάζονται πολλές τεχνικές λεπτομέρειες. Αν είναι δύσκολο να τα καταλάβετε, δεν πειράζει, καθώς τα κύρια συμπεράσματα είναι οι παραδοχές των περιορισμών της τεχνολογίας, η πολυπλοκότητα της διαδικασίας αλληλουχίας και οι πολυάριθμοι τρόποι με τους οποίους εισάγονται λάθη, μεροληψίες και τεχνουργήματα σε κάθε βήμα. Πριν καταδυθούμε στις διάφορες προκλήσεις και τις πιθανές νάρκες που περιμένουν να ανατινάξουν την προσπάθεια αλληλούχισης, ας κάνουμε μια γρήγορη ανάλυση του τι συνεπάγεται ο κατακερματισμός.
Κατακερματισμός του DNA

Αυτή η πρώτη πηγή εξηγεί τι είναι ο κατακερματισμός και παρέχει ένα μικρό πλαίσιο σχετικά με το γιατί χρησιμοποιείται αυτή η ασυνάρτητη διαδικασία κοπής του DNA/RNA σε μικρά θραύσματα προκειμένου να επανασυναρμολογηθεί. Εξηγεί επίσης ένα από τα σημαντικότερα εμπόδια που συχνά εισάγονται κατά τη διάρκεια αυτής της διαδικασίας, γνωστή ως μεροληψία:
Μη τυχαίος κατακερματισμός του DNA στην αλληλούχιση επόμενης γενιάς
"Η τεχνολογία αλληλούχισης επόμενης γενιάς (NGS) βασίζεται στην κοπή του DNA σε μικρά θραύσματα και στη μαζική παράλληλη αλληλούχισή τους. Τα πολλαπλά επικαλυπτόμενα τμήματα που ονομάζονται "reads" συναρμολογούνται σε μια συνεχή αλληλουχία. Για να μειωθούν τα σφάλματα αλληλούχισης, κάθε περιοχή του γονιδιώματος θα πρέπει να αλληλουχηθεί αρκετές δεκάδες φορές. Αυτή η προσέγγιση αλληλούχισης βασίζεται στην υπόθεση ότι τα σπασίματα του γονιδιωματικού DNA είναι τυχαία και ανεξάρτητα από την αλληλουχία".
"Το γονιδίωμα ενός ζωντανού οργανισμού μοιάζει με ένα ράφι γεμάτο βιβλία - χρωμοσώματα που περιέχουν κείμενα αποτελούμενα από γράμματα - νουκλεοτίδια. Οι πρώτες μέθοδοι αποκρυπτογράφησης βιολογικών αλληλουχιών βασίζονταν στην ακριβή αποκοπή ενός συγκεκριμένου τμήματος DNA και στην ακριβή ανάγνωσή του. Μια εναλλακτική και, όπως αρχικά φάνηκε, ασυνάρτητη μέθοδος αλληλούχισης προτάθηκε ήδη από το 19791, σύμφωνα με την οποία τα πολλαπλά αντίγραφα ενός ολόκληρου γονιδιώματος DNA έπρεπε να διασπαστούν σε μικρά θραύσματα, τα οποία αλληλουχήθηκαν και στη συνέχεια οι αλληλουχίες αυτές (που ονομάστηκαν "αναγνώσματα") συναρμολογήθηκαν σε ένα συνεχές κείμενο με βάση τα επικαλυπτόμενα άκρα. Παρ' όλα αυτά, η εκρηκτική ανάπτυξη των αυτοματοποιημένων αλληλουχιών και η πρόοδος στην υπολογιστική ισχύ καθόρισαν τη σημερινή επικράτηση αυτής της μεθόδου, που ονομάζεται τυχαία αλληλούχιση shotgun. Τα σύγχρονα μηχανήματα αλληλούχισης είναι ικανά να διαβάζουν εκατοντάδες εκατομμύρια αναγνώσεις ανά ημέρα, όπου κάθε ανάγνωση αποτελείται από δεκάδες ή εκατοντάδες νουκλεοτίδια.
Το πρώτο βήμα της αλληλούχισης του DNA στην τεχνολογία NGS είναι ο κατακερματισμός του DNA. Τα δείγματα καθαρισμένου DNA διαχωρίζονται σε μικρά θραύσματα, είτε με μηχανικές μεθόδους (π.χ. διάτμηση με υπερήχους και νεφελοποίηση) είτε με ενζυμική πέψη2. Το κατακερματισμένο DNA συνδέεται στα δύο αμβλύ άκρα κάθε θραύσματος με ειδικούς προσαρμογείς, οι οποίοι χρησιμεύουν ως θέσεις πρόσδεσης εκκινητών για την ενίσχυση. Στη συνέχεια, τα συνδεμένα με προσαρμογείς θραύσματα DNA επιλέγονται ως προς το μέγεθος μέσω ηλεκτροφόρησης σε πηκτή αγαρόζης ή με παραμαγνητικά σφαιρίδια- στο στάδιο αυτό αφαιρούνται τα διπλά της σύνδεσης. Στη συνέχεια, τα θραύσματα DNA λιώνουν και τα μονόκλωνα DNA ακινητοποιούνται είτε σε επίπεδες στερεές επιφάνειες μιας κυψέλης ροής (ακολουθιογράφοι Illumina), είτε στην επιφάνεια σφαιριδίων κλίμακας μικρομέτρων (ακολουθιογράφοι 454-Roche και SOLiD), είτε σε ιονισμένες σφαίρες (sequencers (αλληλουχίες) Ion Torrent)3. Η ενίσχυση των προτύπων πραγματοποιείται με PCR σε στερεές επιφάνειες ή με PCR γαλακτώματος σε ξεχωριστούς μικροαντιδραστήρες, σφαιρίδια ή σφαίρες εντός ακολουθιογράφων. Τέλος, η αλληλούχιση επιτυγχάνεται με την ανίχνευση της εκπομπής φωτός ή ιόντων υδρογόνου από κάθε κουκκίδα στη στερεά επιφάνεια ή στις σφαίρες, κατά την ενζυμική σύνδεση συμπληρωματικών νουκλεοτιδίων στις συστάδες πανομοιότυπων μονόκλωνων τμημάτων DNA4.”
Η μεροληψία στα δεδομένα NGS έχει παρατηρηθεί εκτενώς17,18,19,20,21,22,23, αλλά δεν υπάρχει συμφωνία όσον αφορά τις πηγές της παρατηρούμενης μεροληψίας. Έτσι, οι Benjamini και Speed17 ανέφεραν τις κανονικότητες στα μοτίβα μεροληψίας GC και διαπίστωσαν ότι η περιεκτικότητα σε GC επηρεάζει περισσότερο τον αριθμό των θραυσμάτων. Δεδομένου ότι τόσο τα πλούσια σε GC όσο και τα πλούσια σε AT θραύσματα υποεκπροσωπούνταν στα αποτελέσματα της αλληλούχισης, οι συγγραφείς υπέθεσαν ότι η σημαντικότερη αιτία της μεροληψίας GC ήταν η PCR".
"Προτείνουμε να διερευνήσουμε τη μεροληψία που προέρχεται από το στάδιο του κατακερματισμού των διαδικασιών αλληλούχισης NGS. Για πρώτη φορά τολμούμε να δείξουμε ότι η μεροληψία στις αναγνώσεις NGS συσχετίζεται στενά με την μεροληψία που παράγεται από τον ηχοβολισμό καθαρών θραυσμάτων DNA περιορισμού, η οποία αποδείχθηκε ότι είναι ειδική για την αλληλουχία. [27] Ως εκ τούτου, αναφέρουμε και αναλύουμε τη μεροληψία που παράγεται από τρεις μεθόδους θραύσης του DNA, τον ηχοβολισμό, τον νεφελοποίηση και το Covaris και αποδεικνύουμε ότι οι περιπτώσεις μεροληψίας στις διάφορες μεθόδους θραύσης συσχετίζονται σε μεγάλο βαθμό μεταξύ τους και είναι κοινές για όλες τις υδροδυναμικές μεθόδους θραύσης του DNA".
https://www.nature.com/articles/srep04532
Όπως προκύπτει από την πρώτη αυτή πηγή, η αναγκαιότητα κατακερματισμού του DNA σε μικρά κομμάτια για την αλληλουχία τους δημιουργεί το πρόβλημα της εισαγωγής μεροληψίας. Αυτές οι μεροληψίες είναι διαφόρων τύπων και είναι οτιδήποτε προκαλεί παραμόρφωση που μπορεί να ανατρέψει τις γενετικές προβλέψεις και την ακρίβεια του γονιδιώματος. Μπορεί να παρουσιαστεί ως απόκλιση από τις πραγματικές εκτιμήσεις ή ως ετερογένεια (ποικιλομορφία στο περιεχόμενο). Προκειμένου το γονιδίωμα να θεωρείται αξιόπιστο, αυτές οι μεροληψίες πρέπει να λαμβάνονται υπόψη και να διορθώνονται. Χωρίς κάτι τέτοιο, η εργασία θα οδηγήσει σε ανακριβείς ερμηνείες και θα είναι ανεπανάληπτη. Ωστόσο, είναι γνωστό ότι η τεχνολογία, όπως είναι σήμερα, δεν είναι σε θέση να ανταποκριθεί σε αυτή την απαίτηση:
Χαρακτηρισμός και μέτρηση της μεροληψίας σε δεδομένα ακολουθίας
"Η ιδανική αλληλούχιση DNA με κυνηγετικό όπλο όλου του γονιδιώματος θα κατανέμει τις αναγνώσεις ομοιόμορφα σε όλο το γονιδίωμα και χωρίς διακυμάνσεις στην ποιότητα που εξαρτώνται από την αλληλουχία. Όλες οι υπάρχουσες τεχνολογίες αλληλούχισης υπολείπονται αυτού του ιδανικού και παρουσιάζουν διάφορους τύπους και βαθμούς μεροληψίας. Η μεροληψία της αλληλούχισης υποβαθμίζει τις εφαρμογές γονιδιωματικών δεδομένων, συμπεριλαμβανομένης της συναρμολόγησης γονιδιώματος και της ανακάλυψης παραλλαγών, οι οποίες βασίζονται στην κάλυψη σε όλο το γονιδίωμα".
"Η μεροληψία εκδηλώνεται με πολλούς τρόπους. Η μεροληψία κάλυψης είναι μια απόκλιση από την ομοιόμορφη κατανομή των αναγνώσεων σε όλο το γονιδίωμα. Ομοίως, η μεροληψία σφάλματος είναι μια απόκλιση από την προσδοκία ομοιόμορφων ποσοστών αναντιστοιχιών, παρεμβολών και διαγραφών σε αναγνώσεις σε όλο το γονιδίωμα. Το παρόν έγγραφο επικεντρώνεται κυρίως στην μεροληψία κάλυψης επειδή είναι η πιο επιζήμια αποτυχία αλληλούχισης.
Οι τεχνολογίες αλληλούχισης είναι ευάλωτες σε πολλαπλές πηγές μεροληψίας. Οι μέθοδοι που βασίζονται στη βακτηριακή κλωνοποίηση και την αλληλούχιση με τη χημική μέθοδο Sanger [8] υπόκεινται σε πολλές μεροληψίες που μειώνουν την κάλυψη, ιδίως στα ακραία GC, στα παλίνδρομα, στις ανεστραμμένες επαναλήψεις και στις αλληλουχίες που είναι τοξικές για τον βακτηριακό ξενιστή [9-17]. Η αλληλούχιση Illumina [18] έχει αποδειχθεί ότι χάνει την κάλυψη σε περιοχές με υψηλό ή χαμηλό GC [19-22], φαινόμενο που παρατηρείται και σε άλλες τεχνολογίες "επόμενης γενιάς" [3, 6]. Η ενίσχυση με PCR κατά τη διάρκεια της κατασκευής βιβλιοθήκης είναι γνωστή πηγή υποκάλυψης περιοχών με υψηλό GC [20, 21] και παρόμοιες μεροληψίες μπορεί επίσης να εισαχθούν κατά τη διάρκεια της PCR γέφυρας για την ενίσχυση συστάδων στην κυψέλη ροής Illumina [23]. Τα ειδικά σφάλματα της Illumina για τις αλυσίδες μπορούν να οδηγήσουν σε μεροληψίες κάλυψης μειώνοντας την απόδοση του ευθυγραμμιστή [24]. Η Ion Torrent [25], όπως και η 454 [26], χρησιμοποιεί μια χημεία χωρίς τερματιστή, η οποία μπορεί να περιορίζει την ικανότητά της να αλληλουχίζει με ακρίβεια μεγάλα ομοπολυμερή [4, 27, 28], και μπορεί επίσης να είναι ευαίσθητη στις μεροληψίες κάλυψης που εισάγονται από την PCR γαλακτώματος κατά την κατασκευή βιβλιοθήκης. Η Complete Genomics [29] χρησιμοποιεί επίσης ενίσχυση μαζί με μια πολύπλοκη διαδικασία κατασκευής βιβλιοθήκης.
Εκτός από τις πηγές στο υγρό εργαστήριο, η μεροληψία μπορεί να εισαχθεί από οποιοδήποτε από τα υπολογιστικά βήματα στον αγωγό αλληλούχισης. Οι περιορισμοί της επεξεργασίας σήματος και της κλήσης βάσεων θα μπορούσαν να οδηγήσουν σε υποαντιπροσώπευση ή αυξημένα ποσοστά σφάλματος σε ορισμένες θέσεις, όπως και η ανακριβής ευθυγράμμιση. Μια ανακριβής αναφορά ή διαφορές δείγματος-αναφοράς μπορεί να προκαλέσουν διακυμάνσεις στην κάλυψη ή την ακρίβεια που μπορεί να διαγνωστούν λανθασμένα ως μεροληψία αλληλούχισης. Ως εκ τούτου, η ανίχνευση της μεροληψίας είναι μόνο το πρώτο βήμα και πρέπει να ακολουθήσουν πιο λεπτομερή πειράματα για την απόδοση ευθυνών στα στάδια της προετοιμασίας της βιβλιοθήκης, της αλληλούχισης ή του υπολογισμού".
https://genomebiology.biomedcentral.com/articles/10.1186/gb-2013-14-5-r51
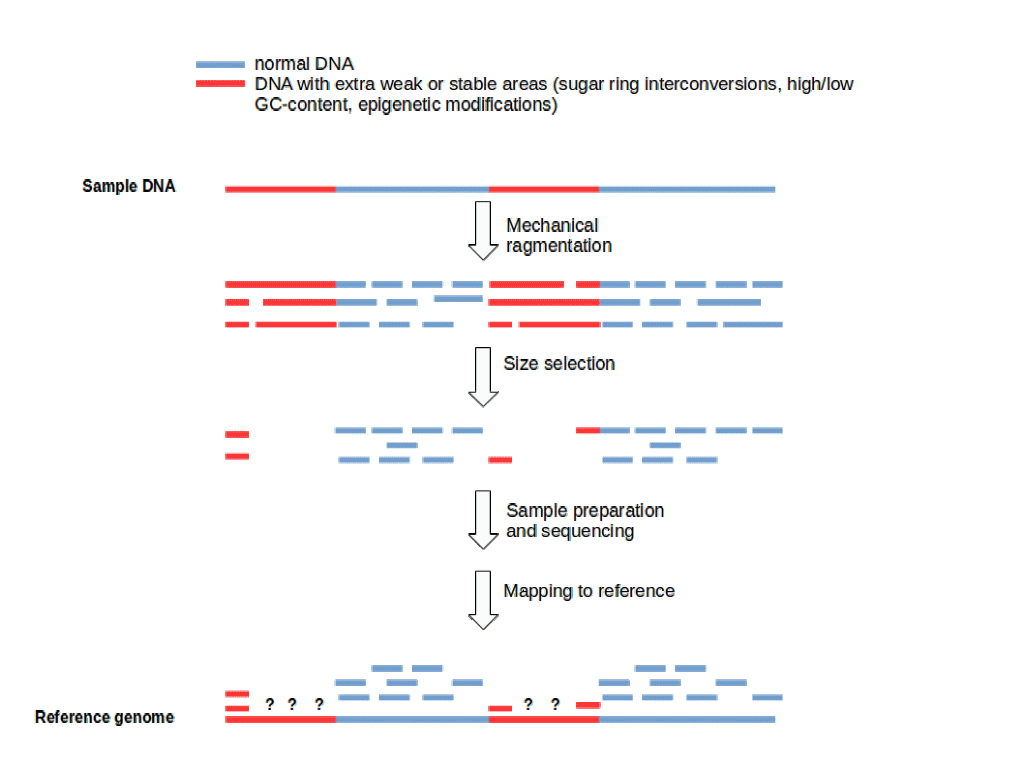
Προφανώς η μεροληψία κατά τη δημιουργία ενός γονιδιώματος θα οδηγούσε σε μια ανακριβή και αναξιόπιστη αναπαράσταση οποιουδήποτε στοιχείου που αλληλουχίζεται. Το πρόβλημα αυτό προέρχεται από το πρώτο κιόλας βήμα της διαδικασίας δημιουργίας του γονιδιώματος. Ο κατακερματισμός είναι κρίσιμος λόγω των τεχνολογικών περιορισμών και δεν μπορεί να αποφευχθεί, οπότε πρέπει να αναμένεται ότι η μεροληψία θα μπει στην εικόνα από την αρχή.
Πώς συμβαίνει αυτός ο κατακερματισμός προκειμένου να δημιουργηθεί η μεροληψία; Αυτή η επόμενη πηγή παρέχει μια καλή επισκόπηση των διαφόρων μεθόδων που χρησιμοποιούνται για τον κατακερματισμό του DNA:
Μέθοδοι Κατακερματισμού του DNA
"Ενώ η προετοιμασία άθικτου DNA υψηλού μοριακού βάρους και η πρόληψη της διάτμησης σε όλα τα περισσότερα στάδια της ροής εργασίας είναι σημαντική, ο κατακερματισμός του DNA είναι ένα απαραίτητο βήμα στην προετοιμασία του δείγματος για τις περισσότερες πλατφόρμες αλληλούχισης. Για παράδειγμα, οι πλατφόρμες αλληλούχισης μικρής ανάγνωσης βασίζονται γενικά σε θραύσματα μήκους ~300-600 bp), ενώ οι πλατφόρμες αλληλούχισης μεγάλης ανάγνωσης είναι συμβατές με θραύσματα μήκους πολλών kb.
Είναι σημαντικό να επιλεγεί μια μέθοδος κατακερματισμού που παράγει αξιόπιστα θραύσματα κατάλληλου μεγέθους χωρίς να εισάγει μεροληψία όσον αφορά τις θέσεις αποκοπής ή τη σύνθεση των βάσεων των θραυσμάτων. Υπάρχουν διάφορες επιλογές για το στάδιο του κατακερματισμού του DNA, η καθεμία με το δικό της μοναδικό προφίλ πλεονεκτημάτων και μειονεκτημάτων, και η επιλογή μιας μεθόδου κατακερματισμού πρέπει να σταθμίζεται προσεκτικά. Οι μέθοδοι που χρησιμοποιούνται περιλαμβάνουν:
Επεξεργασίες με βάση ένζυμα: Αυτές κατακερματίζουν το DNA με ταυτόχρονη διάσπαση και των δύο αλυσίδων ή με τη δημιουργία εγκοπών σε κάθε αλυσίδα dsDNA για την παραγωγή σπασίματος dsDNA. Αυτή η μέθοδος είναι εξαιρετικά ευέλικτη και μπορεί να χρησιμοποιηθεί για τη δημιουργία θραυσμάτων από χαμηλά bp έως πολλά kb σε μήκος.
Ακουστική διάτμηση: Ακουστική ενέργεια μικρού μήκους κύματος και υψηλής συχνότητας εστιάζεται στο δείγμα DNA, διασπώντας φυσικά το μόριο του DNA, χωρίς να απαιτούνται αλλαγές θερμοκρασίας. Η μέθοδος αυτή χρησιμοποιείται για τη δημιουργία θραυσμάτων από χαμηλές εκατοντάδες bp έως πολλά kb σε μήκος.
Ηχοβολισμός: Οι εξειδικευμένοι ηχοβολείς υποβάλλουν το DNA σε μη εστιασμένη ακουστική ενέργεια μεγαλύτερου μήκους κύματος- οι ηχοβολείς απαιτούν περίοδο ψύξης μεταξύ των ριπών ηχοβολισμού. Η μέθοδος αυτή χρησιμοποιείται για τη δημιουργία θραυσμάτων μήκους πολλών kb.
Φυγοκεντρική διάτμηση: Ο ρυθμός φυγοκέντρησης καθορίζει τον βαθμό κατακερματισμού του DNA. Η μέθοδος αυτή χρησιμοποιείται για τη δημιουργία τμημάτων μήκους πολλών kb.
Σημειακή διάτμηση: Μια αντλία σύριγγας παράγει υδροδυναμικές δυνάμεις διάτμησης εντός ενός σωλήνα- το μέγεθος της στένωσης και ο ρυθμός ροής του υγρού καθορίζουν το μέγεθος του θραύσματος DNA. Η μέθοδος αυτή χρησιμοποιείται για τη δημιουργία τμημάτων μήκους πολλών kb.
Διάτμηση με βελόνα: Η επιλογή χαμηλότερης τεχνολογίας βασίζεται στις δυνάμεις διάτμησης που δημιουργούνται με τη διέλευση του DNA μέσω μιας βελόνας μικρού διαμετρήματος. Η μέθοδος αυτή χρησιμοποιείται για τη δημιουργία θραυσμάτων μήκους δεκάδων kb".
https://www.neb.com/applications/ngs-sample-prep-and-target-enrichment/dna-fragmentation
Οι παραπάνω έξι μέθοδοι είναι οι κυριότερες που χρησιμοποιούνται για τον κατακερματισμό του DNA, αλλά όπως συζητήθηκε, υπάρχουν πολλές μεροληψίες και σφάλματα που μπορούν να ενσωματωθούν σε αυτό το κρίσιμο βήμα και τα οποία θα μεταφερθούν στα επόμενα βήματα που απαιτούνται για την αλληλούχιση του γονιδιώματος. Αυτό δημιουργεί αποτελέσματα αλληλούχισης που δεν είναι επαναλήψιμα και μη αναπαραγώγιμα. Οι περιορισμοί και οι δυσκολίες που συναντώνται κατά τη διάτμηση/κατακερματισμό, οι οποίες μπορούν να οδηγήσουν σε αυτά τα προβλήματα, περιγράφονται συνοπτικά εδώ:
Πώς λειτουργεί: Ακριβής Διάτμηση του χρωμοσωμικού DNA
"Πρόβλημα: Όλες οι σύγχρονες, καθώς και οι μεθοδολογίες αλληλούχισης επόμενης και τρίτης γενιάς εξαρτώνται από την παραγωγή τμημάτων DNA από αρχικά μόρια χρωμοσωμικού DNA μήκους MegaBase. Οι γενικές απαιτήσεις είναι ότι τα θραύσματα αυτά πρέπει να είναι τυχαία και παρόμοιου μεγέθους. Έχουν αναπτυχθεί πολλές μέθοδοι κατακερματισμού του DNA. Οι περισσότερες από αυτές τις μεθόδους περιλαμβάνουν περιορισμούς και δυσκολίες, όπως υψηλό κόστος, κατακερματισμό, ευρεία κατανομή μεγέθους θραυσμάτων ή μη αναστρέψιμες βλάβες στα θραύσματα DNA. Επιπλέον, τα αποτελέσματα μπορεί να εξαρτώνται από τη συγκέντρωση και/ή να μην είναι ιδιαίτερα αναπαραγώγιμα".
https://www.google.com/amp/s/www.labmanager.com/how-it-works/accurate-shearing-of-chromosomal-dna-19660/amp

Αυτή η επόμενη πηγή διαφοροποιεί μεταξύ ενός φυσικού μέσου κατακερματισμού με υπερηχητικό, το οποίο είναι γνωστό ότι προκαλεί απώλεια DNA, και του ενζυμικού κατακερματισμού, ο οποίος δεν έχει αξιολογηθεί προσεκτικά και οδηγεί σε τεχνάσματα και μεροληψίες. Αναφέρει ότι η διαδικασία αλληλούχισης περιλαμβάνει πολλαπλά βήματα για τα οποία μπορούν να εισαχθούν μεροληψίες και σφάλματα αλληλούχισης. Παρόλο που έχουν γίνει προσπάθειες για την ελαχιστοποίηση των σφαλμάτων, αυτά εξακολουθούν να υφίστανται. Υποστηρίζεται ότι λόγω της επιμονής αυτών των σφαλμάτων και των τεχνουργημάτων, μπορεί να εκληφθούν ως μεταλλάξεις και να προστεθούν στο γονιδίωμα.
Τεχνούργημα αλληλούχισης που προέρχεται από μια μέθοδο προετοιμασίας βιβλιοθήκης με χρήση ενζυμικού κατακερματισμού
"Ο κατακερματισμός του DNA είναι ένα θεμελιώδες βήμα κατά την προετοιμασία της βιβλιοθήκης στην αλληλούχιση μικρής ανάγνωσης με βάση τη σύλληψη υβριδισμού. Μέχρι στιγμής έχει χρησιμοποιηθεί ο υπερηχοτονισμός για την προετοιμασία DNA κατάλληλου μεγέθους, αλλά η μέθοδος αυτή συνδέεται με σημαντική απώλεια δείγματος DNA. Πιο πρόσφατα, σε μελέτες έχουν χρησιμοποιηθεί μέθοδοι προετοιμασίας βιβλιοθήκης που βασίζονται στον ενζυμικό κατακερματισμό με ενδονουκλεάσες DNA για την ελαχιστοποίηση της απώλειας DNA, ιδίως σε δείγματα νανο-ποσοτήτων. Ωστόσο, παρά την ευρεία χρήση τους, η επίδραση του ενζυμικού κατακερματισμού στις προκύπτουσες αλληλουχίες δεν έχει αξιολογηθεί προσεκτικά. Εδώ, χρησιμοποιήσαμε συγκρίσεις ανά ζεύγη σωματικών παραλλαγών των ίδιων δειγμάτων DNA όγκου που παρασκευάστηκαν με τη χρήση μεθόδων υπερήχων και ενζυμικού κατακερματισμού. Η ανάλυσή μας αποκάλυψε σημαντικά μεγαλύτερο αριθμό επαναλαμβανόμενων τεχνητών SNVs/indels στις βιβλιοθήκες που έχουν υποστεί επεξεργασία με ενδονουκλεάση σε σύγκριση με εκείνες που δημιουργήθηκαν με υπερήχους. Αυτά τα τεχνουργήματα χαρακτηρίζονταν από παλίνδρομη δομή στο γονιδιωματικό πλαίσιο, μεροληψία θέσης στις αλληλουχημένες αναγνώσεις και υποκαταστάσεις πολλών νουκλεοτιδίων".
"Η αλληλούχιση γονιδιώματος με τη χρήση σύλληψης υβριδισμού περιλαμβάνει πολλαπλά στάδια, συμπεριλαμβανομένης της επεξεργασίας ιστού, της αποθήκευσης ιστού, της απομόνωσης DNA, του κατακερματισμού DNA, του υβριδισμού ανιχνευτών, της ενίσχυσης βιβλιοθήκης, της αλληλούχισης και της ανάλυσης πληροφορικής [2–5].. Σφάλματα αλληλούχισης μπορούν να εισαχθούν σε οποιοδήποτε από αυτά τα στάδια και τα νουκλεοτίδια μπορούν να τροποποιηθούν περαιτέρω μέσω οξείδωσης κατά την επεξεργασία ιστών, την αποθήκευση ιστών, την απομόνωση του DNA και τον κατακερματισμό του DNA [2–4, 6]. Τα σφάλματα ενσωμάτωσης νουκλεοτιδίων μπορούν με τη σειρά τους να δημιουργήσουν μεροληψίες στην αντίδραση πολυμεράσης, να επηρεάσουν την ακριβή ενίσχυση της βιβλιοθήκης και να δημιουργήσουν θόρυβο αλληλούχισης [2–4]. Αν και έχουν καταβληθεί σημαντικές προσπάθειες για την ελαχιστοποίηση αυτού του θορύβου αλληλούχισης πειραματικά, τα στοχαστικά σφάλματα παραμένουν επίμονα [2–4].
Ο υπερηχοτονισμός αποτελεί εδώ και πολύ καιρό μια τυπική μέθοδο για τον κατακερματισμό του DNA κατά την αλληλούχιση μικρής ανάγνωσης με βάση τη σύλληψη υβριδισμού. Ο υπερηχητισμός δημιουργεί ομοιόμορφες τομές στο DNA σε ολόκληρο το γονιδίωμα, παρέχοντας έτσι ένα απλό μέσο ελέγχου του μεγέθους των θραυσμάτων με μη προκατειλημμένο τρόπο [2, 3].. Ωστόσο, η φυσική διασπορά του διαλύματος DNA κατά τη διαδικασία οδηγεί συχνά σε απώλεια δείγματος DNA, η οποία μπορεί να είναι κρίσιμη όταν η ποσότητα του δείγματος περιορίζεται σε ποσότητες νανο- ή πικογραμμαρίων, όπως συμβαίνει με τα θραύσματα βιοψίας ιστού. Διατίθενται διάφορα εμπορικά κιτ προετοιμασίας βιβλιοθήκης, συμπεριλαμβανομένων των κιτ HyperPlus (KAPA Biosystems), SureSelect QXT (Agilent Technologies), Fragmentase (New England Biolabs) και Nextera Tagmentation (Illumina), καθένα από τα οποία χρησιμοποιεί ενδονουκλεάσες ή τρανσποζάσες για τον κατακερματισμό του DNA. Παρόλο που αυτά τα κιτ ελαχιστοποιούν την απώλεια DNA, παραμένει σε μεγάλο βαθμό άγνωστο σε ποιο βαθμό τα σφάλματα αλληλούχισης προκαλούνται από τη διαδικασία ενζυμικού κατακερματισμού".
"Ο κατακερματισμός του DNA είναι ένα απαραίτητο βήμα στην προετοιμασία των νουκλεϊκών οξέων, καθώς η ποιότητα της αλληλούχισης εξαρτάται τόσο από την τυχαιότητα του κατακερματισμού του DNA όσο και από την επικάλυψη των θραυσμάτων που προκύπτουν. Επιπλέον, επειδή το μέγεθος των θραυσμάτων τείνει να διαφέρει μεταξύ των πλατφορμών NGS και των τμημάτων αλληλούχισης, ο αποτελεσματικός έλεγχος του μεγέθους των θραυσμάτων DNA είναι επιτακτικός".
"Δεδομένου ότι οι ίδιες οι ενδονουκλεάσες είναι ανίκανες να ενσωματώσουν νουκλεοτίδια στο DNA ή να προκαλέσουν μεταλλάξεις [19], υποθέτουμε ότι οι μεταλλάξεις προκύπτουν μετά από ενζυμικό κατακερματισμό κατά τη διάρκεια της "διαδικασίας συμπλήρωσης" που ενορχηστρώνεται από την πολυμεράση DNA για την επιδιόρθωση του άκρου ("End repair & A-tailing enzyme" πριν από τη σύνδεση προσαρμογέα στο κιτ HyperPlus). Ο υπερηχητικός συντονισμός διασπά τυχαία μόρια DNA σε διαφορετικές γονιδιωματικές θέσεις και, ως εκ τούτου, κατά την επακόλουθη διαδικασία πλήρωσης, νουκλεοτίδια ενσωματώνονται σε διαφορετικές γονιδιωματικές θέσεις σε διαφορετικά μόρια DNA. Ακόμη και αν ένα λανθασμένο νουκλεοτίδιο ενσωματωθεί στις θέσεις που διασπάστηκαν, το προκύπτον τεχνούργημα δεν θα αναγνωριζόταν ως μετάλλαξη, επειδή δεν θα εμφανιζόταν σταθερά στην ίδια θέση σε διαφορετικά μόρια. Ωστόσο, επειδή η ενδονουκλεάση HyperPlus κόβει κατά προτίμηση συγκεκριμένες θέσεις στο DNA, όταν ενσωματώνεται ένα λανθασμένο νουκλεοτίδιο, το προκύπτον τεχνούργημα θα μπορούσε να αναγνωριστεί λανθασμένα ως μετάλλαξη, επειδή εμφανίζεται επανειλημμένα στην ίδια θέση σε διαφορετικά μόρια".
"Βρήκαμε έναν σημαντικό αριθμό σωματικών SNVs/indels στην ανάλυση ζεύγους των έξι δειγμάτων όγκου με τη χρήση της επεξεργασίας SureSelect για τα φυσιολογικά δείγματα και της επεξεργασίας HyperPlus για τα δείγματα όγκου (SH). Θεωρήσαμε ότι αυτός ο θόρυβος θα μπορούσε να αποφευχθεί με τη χρήση της ίδιας μεθόδου κατακερματισμού του DNA για τα ζευγαρωμένα δείγματα (δηλαδή συνδυασμός HH) και το δοκιμάσαμε αυτό χρησιμοποιώντας δείγματα από δύο περιπτώσεις καρκίνου του ορθού. Παρόλο που επιβεβαιώσαμε μια σημαντική μείωση του αριθμού των SNVs/indels με τη χρήση μόνο μιας μεθόδου κατακερματισμού, κατά την προσεκτική εξέταση, εντοπίσαμε την παραμονή του θορύβου HyperPlus μεταξύ των SNVs/indels που προέκυψαν από τον συνδυασμό HH- ο θόρυβος αυτός ταξινομήθηκε συχνά από τον αλγόριθμο σε άλλες συγκρίσεις ανά ζεύγη και χαρακτηριζόταν από παλίνδρομη δομή. Το εύρημα αυτό ενισχύει την πρότασή μας για τον κίνδυνο τα επίμονα σφάλματα να συγχέονται με γνήσιες μεταλλάξεις λόγω της επαναλαμβανόμενης εμφάνισής τους σε μια κοόρτη".
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0227427
Κατακερματισμός RNA

Οι παραπάνω μέθοδοι περιέγραψαν λεπτομερώς τη διαδικασία κατακερματισμού όπως αυτή σχετίζεται με το DNA. Όταν έχουμε να κάνουμε με RNA, υπάρχουν πρόσθετα βήματα που πρέπει να γίνουν προκειμένου να μπορέσουμε να αλληλουχήσουμε ένα γονιδίωμα. Αυτό γίνεται με δύο διαφορετικούς τρόπους. Η πρώτη μέθοδος επιτυγχάνεται με τη λήψη RNA και την κάλυψή του σε cDNA, το οποίο είναι DNA που συντίθεται από ένα πρότυπο RNA σε μια αντίδραση που καταλύεται από το ένζυμο αντίστροφη μεταγραφάση και πραγματοποιείται μέσω RT-PCR. Στη συνέχεια, χρησιμοποιούνται οι ίδιες μέθοδοι διάτμησης του DNA που αναφέρθηκαν παραπάνω για τη διάτμηση του cDNA ώστε να δημιουργηθούν τα απαραίτητα θραύσματα για την αλληλούχιση.
Η δεύτερη μέθοδος είναι μια χημική διαδικασία κατακερματισμού που χρησιμοποιεί ιόντα μετάλλων όπως Mg++ και Zn++ σε υψηλές θερμοκρασίες και αλκαλικές συνθήκες ή ενζυμικός κατακερματισμός με χρήση RNάσης. Αυτή η διαδικασία αλληλούχισης RNA περιγράφεται λεπτομερώς σε αυτή την επόμενη πηγή και δείχνει πώς η εξαιρετικά περίπλοκη ροή εργασίας για την αλληλούχιση RNA παράγει εύκολα σφάλματα. Αυτό προφανώς οδηγεί σε ανακριβείς και λανθασμένες ερμηνείες:
Μεροληψία στην προετοιμασία βιβλιοθήκης RNA-seq: Τρέχουσες προκλήσεις και λύσεις
"Αν και η αλληλούχιση RNA (RNA-seq) έχει γίνει η πιο προηγμένη τεχνολογία για την ανάλυση του μεταγραφώματος, αντιμετωπίζει επίσης διάφορες προκλήσεις. Όπως όλοι γνωρίζουμε, η ροή εργασίας του RNA-seq είναι εξαιρετικά περίπλοκη και είναι εύκολο να παραχθεί μεροληψία. Αυτό μπορεί να βλάψει την ποιότητα του συνόλου δεδομένων RNA-seq και να οδηγήσει σε εσφαλμένη ερμηνεία του αποτελέσματος της αλληλούχισης. Συνεπώς, η λεπτομερής κατανόηση της πηγής και της φύσης αυτών των μεροληψιών είναι απαραίτητη για την ερμηνεία των δεδομένων RNA-seq, την εξεύρεση μεθόδων για τη βελτίωση της ποιότητας των πειραματικών RNA-seq ή την ανάπτυξη εργαλείων βιοπληροφορικής για την αντιστάθμιση αυτών των μεροληψιών. Εδώ, συζητάμε τις πηγές των πειραματικών μεροληψιών στο RNA-seq".
"Ωστόσο, το RNA-seq είναι μια διαδικασία εξαιρετικά περίπλοκη, που περιλαμβάνει την εξαγωγή και τον καθαρισμό RNA, την κατασκευή βιβλιοθήκης, την αλληλούχιση και την ανάλυση βιοπληροφορικής. Αυτές οι διαδικασίες μπορούν αναπόφευκτα να εισάγουν ορισμένες αποκλίσεις (Πίνακας 1), οι οποίες επηρεάζουν την ποιότητα των συνόλων δεδομένων RNA-seq και οδηγούν σε εσφαλμένη ερμηνεία τους. Ως εκ τούτου, η κατανόηση αυτών των μεροληψιών είναι ζωτικής σημασίας για την αποφυγή λανθασμένης ερμηνείας των δεδομένων και για την αξιοποίηση όλων των δυνατοτήτων αυτής της ισχυρής τεχνολογίας".
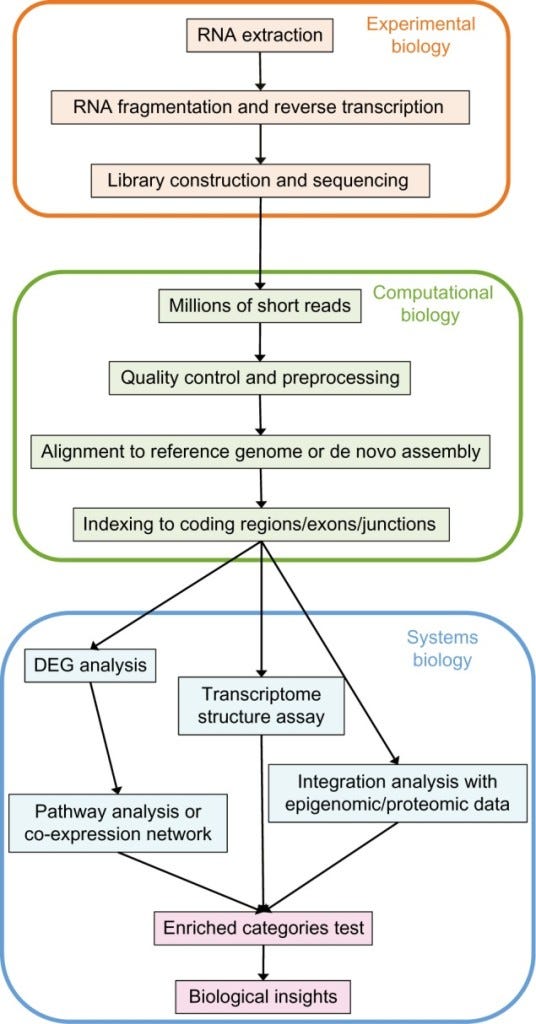
"Σε γενικές γραμμές, η αντιπροσωπευτική ροή εργασίας της ανάλυσης RNA-seq περιλαμβάνει την εξαγωγή και τον καθαρισμό του RNA από κύτταρο ή ιστό, την προετοιμασία της βιβλιοθήκης αλληλούχισης, συμπεριλαμβανομένης της κατακερματισμού, της γραμμικής ενίσχυσης ή της ενίσχυσης με PCR, την αλληλούχιση RNA και την επεξεργασία και ανάλυση των δεδομένων αλληλούχισης (Εικόνα 1). Οι κοινώς χρησιμοποιούμενες πλατφόρμες NGS, συμπεριλαμβανομένων των Illumina και Pacific Biosciences, χρειάζονται ενίσχυση με PCR κατά τη διάρκεια της κατασκευής βιβλιοθήκης για να αυξηθεί ο αριθμός των μορίων cDNA ώστε να καλυφθούν οι ανάγκες της αλληλούχισης. Παρ' όλα αυτά, το πιο προβληματικό βήμα στις διαδικασίες προετοιμασίας δειγμάτων είναι η ενίσχυση. Αυτό οφείλεται στο γεγονός ότι η ενίσχυση με PCR εισάγει στοχαστικές μεροληψίες, οι οποίες μπορούν να μεταδοθούν σε μεταγενέστερους κύκλους [2].Επιπλέον, η PCR ενισχύει επίσης διαφορετικά μόρια με άνισες πιθανότητες, οδηγώντας στην άνιση ενίσχυση των μορίων cDNA [3, 4].Πρόσφατα, οι ερευνητές πρότειναν διάφορες διαφορετικές μεθόδους προκειμένου να μειώσουν την ενίσχυση με PCR, όπως πρωτόκολλα χωρίς PCR και ισοθερμική ενίσχυση. Παρ' όλα αυτά, οι μέθοδοι αυτές δεν είναι τέλειες και εξακολουθούν να παρουσιάζουν ορισμένα τεχνουργήματα και μεροληψίες της αλληλούχισης. Κατά συνέπεια, η κατανόηση αυτών των μεροληψιών είναι ζωτικής σημασίας για την απόκτηση αξιόπιστων δεδομένων και θα παράσχει ορισμένες χρήσιμες συμβουλές στον ερευνητή."
3.3. Κατακερματισμός RNA
"Επί του παρόντος, το RNA είναι συνήθως κατακερματισμένο λόγω του περιορισμού του μήκους ανάγνωσης (<600 bp) των τεχνολογιών αλληλούχισης και της ευαισθησίας της ενίσχυσης σε μεγάλα μόρια cDNA. Υπάρχουν δύο κύριες προσεγγίσεις κατακερματισμού του RNA: η χημική (με χρήση ιόντων μετάλλων) και η ενζυμική (με χρήση RNase III) [30]. Συνήθως, ο κατακερματισμός του RNA γίνεται με τη χρήση ιόντων μετάλλων, όπως Mg++ και Zn++, σε υψηλές θερμοκρασίες και αλκαλικές συνθήκες. Αυτή η μέθοδος αποδίδει ακριβέστερη ταυτοποίηση μεταγραφών από την πέψη με RNάση ΙΙΙ [31]. Το αποτέλεσμα αυτό επιβεβαιώθηκε επίσης στους Wery et al. [31]. Επιπλέον, τα άθικτα RNA μπορούν να μεταγραφούν αντίστροφα (RT) σε cDNA με αντίστροφη μεταγραφάση, στη συνέχεια κατακερματισμένα. Στη συνέχεια, το cDNA κατακερματίστηκε με τη χρήση ενζυμικής ή φυσικής μεθόδου. Παραδείγματα της ενζυμικής μεθόδου είναι η πέψη με DNase I, η μη ειδική ενδονουκλεάση (όπως η NEBNext dsDNA Fragmentase της New England Biolabs) και ο κατακερματισμός DNA με τη μεσολάβηση τρανσποζάσης (Illumina Nextera XT). Ωστόσο, η μέθοδος της τρανσποζάσης Tn5 έδειξε μεροληψία ως προς την αλληλουχία [32], η οποία είναι η προτιμώμενη μέθοδος όταν είναι διαθέσιμες μόνο μικρές ποσότητες cDNA, δεδομένου ότι ο κατακερματισμός του cDNA και η σύνδεση του προσαρμογέα συνδέονται σε ένα βήμα [33]. Μελέτες έχουν δείξει ότι οι μη ειδικές ενδονουκλεάσες περιορισμού δείχνουν μικρότερη μεροληψία αλληλουχίας και έχει αποδειχθεί ότι έχουν παρόμοιες επιδόσεις με τις φυσικές μεθόδους όσον αφορά την μεροληψία αλληλουχίας της θέσης διάσπασης και την ομοιομορφία της κάλυψης του DNA-στόχου [34, 35]. Ένα άλλο πλεονέκτημα της ενζυμικής μεθόδου είναι ότι είναι εύκολο να αυτοματοποιηθεί [36]. Η φυσική μέθοδος περιλαμβάνει την ακουστική διάτμηση, τον ηχητικό καθαρισμό και την υδροδυναμική [17, 37, 38], οι οποίες μπορούν επίσης να παρουσιάσουν μη τυχαία μεροληψία κατακερματισμού του DNA [35]. Ωστόσο, η φυσική μέθοδος κατακερματισμού cDNA είναι λιγότερο επιδεκτική αυτοματοποίησης από ό,τι ο κατακερματισμός RNA. Ως εκ τούτου, η φυσική μέθοδος θα αντικατασταθεί από τα εμπορικά διαθέσιμα κιτ και την ενζυμική μέθοδο".
5. Συζήτηση και Συμπεράσματα
"Επί του παρόντος, το RNA-seq χρησιμοποιείται ευρέως στη βιολογική, ιατρική, κλινική και φαρμακευτική έρευνα. Ωστόσο, όλες αυτές οι μελέτες αλληλούχισης περιορίζονται από την ακρίβεια των υποκείμενων πειραμάτων αλληλούχισης, επειδή η τεχνολογία RNA-seq μπορεί να εισάγει διάφορα σφάλματα και μεροληψίες στην προετοιμασία του δείγματος, την κατασκευή βιβλιοθήκης, την αλληλούχιση και την απεικόνιση κ.λπ.".
https://www.hindawi.com/journals/bmri/2021/6647597/

Τα ίδια προβλήματα όσον αφορά τα σφάλματα και τις μεροληψίες που σχετίζονται με τον κατακερματισμό του DNA εξακολουθούν να εμφανίζονται με τον κατακερματισμό του cDNA που μετατρέπεται από RNA. Αυτό περιπλέκεται επίσης περαιτέρω από τα σφάλματα/την επιμόλυνση που εμπλέκονται στη διαδικασία RT-PCR που χρησιμοποιείται για τη σύνθεση του cDNA, για την οποία θα αναφερθώ σε άλλη χρονική στιγμή. Ωστόσο, ακόμη και χωρίς αυτό το βήμα και με τη χρήση μεταλλικών ιόντων/RNases απευθείας στο RNA, εξακολουθούν να εμφανίζονται προβλήματα:
Αρχές διάσπασης Νουκλεϊκών Οξέων από Μεταλλικά Ιόντα
"Συχνά οι μοριακοί βιολόγοι που ασχολούνται με το DNA και το RNA πιστεύουν ότι οι ιδιότητες των μεταλλικών ιόντων είναι απλούστερες από τις ιδιότητες των νουκλεϊκών οξέων λόγω του μικρότερου μεγέθους τους. Στην πραγματικότητα, τόσο τα νουκλεϊκά οξέα όσο και τα μεταλλικά ιόντα παρουσιάζουν σημαντική πολυπλοκότητα στις αλληλεπιδράσεις τους και οι αλληλεπιδράσεις αυτές θα μπορούσαν να επηρεάσουν τις χημικές και βιοχημικές ιδιότητες και των δύο μερών.
Τα μεταλλικά ιόντα απαιτούνται συνήθως για την προώθηση και τη σταθεροποίηση λειτουργικά ενεργών ή εγγενών διαμορφώσεων των νουκλεϊκών οξέων, αλλά μπορούν επίσης να παγιδεύσουν πολυνουκλεοτίδια σε ανενεργές διαμορφώσεις (Heilman-Miller et al. 2001). Εκτός από τους δομικούς ρόλους, τα περισσότερα πολυδύναμα μεταλλικά ιόντα (M22+) μπορούν να προκαλέσουν διάσπαση (δηλαδή θραύση, διάσπαση, κατακερματισμό, αποπολυμερισμό ή ρήξη) των νουκλεϊκών οξέων. Οι αντιδράσεις αυτές μπορεί να είναι είτε μη ειδικές είτε να εξαρτώνται από τη χημική φύση των νουκλεοτιδικών καταλοίπων, την αλληλουχία του νουκλεϊκού οξέος ή τη δευτεροταγή ή/και τριτοταγή δομή. Η εξειδίκευση αυτών των αντιδράσεων εξαρτάται τόσο από τη διαμόρφωση του νουκλεϊκού οξέος και τους τρόπους πρόσδεσης των μετάλλων όσο και από τις ιδιότητες των ιόντων μετάλλων".
"Εδώ, επανεξετάζουμε κριτικά τους θεμελιώδεις παράγοντες που επηρεάζουν την αποτελεσματικότητα και την εξειδίκευση της διάσπασης νουκλεϊκών οξέων που προάγεται από ιόντα μετάλλων. Αυτοί οι (συχνά αγνοούμενοι) παράγοντες μπορεί να έχουν δραματικές επιπτώσεις στα πειράματα ή μπορεί να οδηγήσουν σε τεχνουργήματα ή παρερμηνεία των πειραματικών δεδομένων."
"Το DNA και το RNA διαφέρουν έντονα ως προς τη σταθερότητα των φωσφοδιεστερικών δεσμών τους σε υδατικά διαλύματα. Γενικά, το DNA θεωρείται πιο σταθερό από το RNA (Li και Breaker 1999- Thorp 2000- Williams κ.ά. 1999). Πράγματι, το RNA διασπάται εύκολα ακόμη και σε ήπια αλκαλικά διαλύματα, ενώ το DNA είναι σταθερό υπό αυτές τις συνθήκες. Η μόλυνση των διαλυμάτων από RNάσες και ιόντα Μ2+ μπορεί να οδηγήσει σε γρήγορη αποικοδόμηση του RNA ακόμη και σε ουδέτερο pH και χαμηλές θερμοκρασίες".
https://www.google.com/url?sa=t&source=web&rct=j&url=https://static1.squarespace.com/static/5ae2118f697a984bb60bd6c3/t/5b2ac74c352f53d7dbf5609e/1529530193348/Dallas_2004.pdf&ved=2ahUKEwj-s9bvz9PxAhVba80KHT_CDCwQFjAAegQIBRAC&usg=AOvVaw1EEoxCIULXXuu8KMeKMldg

Αυτές οι τελευταίες πηγές παρέχουν περισσότερες λεπτομέρειες σχετικά με τους διάφορους τύπους μεροληψιών και σφαλμάτων που μπορούν να εισαχθούν κατά τη διαδικασία κατακερματισμού του RNA. Σε αυτά περιλαμβάνονται ανακριβή στατιστικά μοντέλα, μεροληψίες θέσης λόγω της αποικοδόμησης του RNA ή/και του βήματος ενίσχυσης της PCR, μεροληψίες συγκεκριμένης αλληλουχίας που βασίζονται σε εσφαλμένες υποθέσεις και η αβεβαιότητα της βιοχημείας πολλών από τα εμπλεκόμενα βήματα. Όπως και στον κατακερματισμό του DNA, η μόλυνση, τα σφάλματα, τα τεχνουργήματα και οι μεροληψίες είναι κοινά και στον κατακερματισμό του RNA:
Τα μοντέλα μίξης αποκαλύπτουν πολλαπλούς τύπους μεροληψίας θέσης σε δεδομένα RNA-Seq και οδηγούν σε ακριβείς εκτιμήσεις της συγκέντρωσης μεταγραφών
"Παρά τα πλεονεκτήματα αυτά, η λήψη ακριβών μετρήσεων ποσοτικοποίησης μεταγράφων από το RNA-Seq έχει αποδειχθεί δύσκολη. Ένας από τους κύριους λόγους της ανακρίβειας είναι η αποτυχία των στατιστικών μοντέλων που χρησιμοποιούνται για την εξαγωγή των μετρήσεων να αναπαραστήσουν σωστά τις μεροληψίες που είναι εγγενείς στα δεδομένα RNA-Seq. Το στατιστικό μοντέλο της αρχικής έκδοσης του Cufflinks [2], για παράδειγμα, υποθέτει ότι τα θραύσματα cDNA που παράγονται από το RNA-Seq κατανέμονται ομοιόμορφα κατά μήκος των μεταγράφων. Στην πραγματικότητα, ωστόσο, αυτή η υπόθεση σπάνια πληρούται και, ως εκ τούτου, οι μετρήσεις ποσοτικοποίησης με αυτή την έκδοση του Cufflinks είναι συχνά ανακριβείς.
Ένας τύπος μεροληψίας που επηρεάζει την ποσοτικοποίηση μεταγραφών από δεδομένα RNA-Seq είναι το αποτέλεσμα της προτίμησης του κατακερματισμού, δηλαδή της διαδικασίας που παράγει θραύσματα cDNA από μεταγραφή RNA, να παράγει θραύσματα σε ορισμένες θέσεις εντός της μεταγραφής, π.χ. στην αρχή ή/και στο τέλος της μεταγραφής [3]. Ως εκ τούτου, αυτός ο τύπος μεροληψίας αναφέρεται ως μεροληψία θέσης [4]. Η μεροληψία θέσης μπορεί επίσης να προκληθεί από μεροληψία στο ίδιο το RNA, για παράδειγμα, λόγω αποικοδόμησης του RNA που έχει ως αποτέλεσμα τη συντόμευση του RNA. Ένα άλλο είδος μεροληψίας στο RNA-Seq εισάγεται κατά τη διάρκεια της σύνδεσης, της ενίσχυσης και της αλληλούχισης NGS [5]. Αυτή η μεροληψία συσχετίζεται με την αλληλουχία RNA ενός μεταγράφου και ως εκ τούτου ονομάζεται μεροληψία συγκεκριμένης αλληλουχίας [4]".
https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1005515#sec001
Βελτίωση των εκτιμήσεων έκφρασης RNA-Seq με διόρθωση της μεροληψίας των θραυσμάτων
"Δυστυχώς, οι σημερινοί τεχνολογικοί περιορισμοί των sequencers απαιτούν τα μόρια cDNA να αντιπροσωπεύουν μόνο μερικά θραύσματα του RNA που εξετάζεται. Τα θραύσματα cDNA λαμβάνονται με μια σειρά βημάτων, συχνά συμπεριλαμβανομένης της αντίστροφης μεταγραφής που προετοιμάζεται με τυχαία εξαμερή (RH), ή με ολιγο(dT). Τα περισσότερα πρωτόκολλα περιλαμβάνουν επίσης ένα στάδιο κατακερματισμού, συνήθως υδρόλυση RNA ή νεφελοποίηση, ή εναλλακτικά κατακερματισμό cDNA με επεξεργασία DNase I ή ηχοβολισμό. Πολλές τεχνολογίες αλληλούχισης απαιτούν επίσης περιορισμένα μήκη cDNA, οπότε μπορεί να περιλαμβάνεται ένα τελικό βήμα κοπής σε πηκτή για την επιλογή μεγέθους. "
"Η τυχαιότητα που ενυπάρχει σε πολλά από τα βήματα προετοιμασίας για RNA-Seq οδηγεί σε θραύσματα των οποίων τα σημεία εκκίνησης (σε σχέση με τα μεταγράμματα από τα οποία αλληλουχήθηκαν) φαίνεται να επιλέγονται περίπου ομοιόμορφα τυχαία. Η παρατήρηση αυτή αποτέλεσε τη βάση των παραδοχών που διέπουν ορισμένες προσεγγίσεις ανάλυσης RNA-Seq οι οποίες, με όρους επιστήμης της πληροφορικής, αντιστρέφουν την "αναγωγή" της εκτίμησης του μεταγραφώματος στην αλληλούχιση DNA [2-6]. Ωστόσο, η πρόσφατη προσεκτική ανάλυση αποκάλυψε τόσο τις μεροληψίες θέσης [7] όσο και τις μεροληψίες συγκεκριμένης αλληλουχίας [8, 9] στα αλληλουχημένα θραύσματα. Η μεροληψία θέσης αναφέρεται σε ένα τοπικό φαινόμενο κατά το οποίο τα θραύσματα εντοπίζονται κατά προτίμηση είτε προς την αρχή είτε προς το τέλος των μεταγράφων. Η μεροληψία που σχετίζεται με την αλληλουχία είναι ένα συνολικό φαινόμενο όπου η αλληλουχία που περιβάλλει την αρχή ή το τέλος των πιθανών θραυσμάτων επηρεάζει την πιθανότητα επιλογής τους για αλληλούχιση. Αυτές οι μεροληψίες μπορούν να επηρεάσουν τις εκτιμήσεις έκφρασης [10] και, ως εκ τούτου, είναι σημαντικό να διορθώνονται κατά τη διάρκεια της ανάλυσης RNA-Seq.
Παρόλο που πολλές μεροληψίες μπορούν να αποδοθούν σε ιδιαιτερότητες των πρωτοκόλλων προετοιμασίας (βλ. Εικόνα 2 και [8]), δεν είναι επί του παρόντος δυνατό να προβλεφθούν οι κατανομές θραυσμάτων απευθείας από ένα πρωτόκολλο. Αυτό οφείλεται σε πολλούς παράγοντες, συμπεριλαμβανομένης της αβεβαιότητας στη βιοχημεία πολλών βημάτων και του άγνωστου σχήματος και της επίδρασης της δευτεροταγούς δομής του RNA σε ορισμένες διαδικασίες [10]".
https://genomebiology.biomedcentral.com/articles/10.1186/gb-2011-12-3-r22

Συνοψίζοντας:
Η τεχνολογία αλληλούχισης επόμενης γενιάς (NGS) βασίζεται στην κοπή του DNA σε μικρά θραύσματα και στη μαζική παράλληλη αλληλούχισή τους
Αυτή η προσέγγιση αλληλούχισης βασίζεται στην υπόθεση ότι τα σπασίματα του γονιδιωματικού DNA είναι τυχαία και ανεξάρτητα από την αλληλουχία
Μια εναλλακτική και, όπως αρχικά φάνηκε, ασυνάρτητη μέθοδος αλληλούχισης προτάθηκε το 1979, σύμφωνα με την οποία τα πολλαπλά αντίγραφα ενός ολόκληρου γονιδιώματος DNA έπρεπε να διασπαστούν σε μικρά θραύσματα, τα οποία αλληλουχήθηκαν και στη συνέχεια αυτές οι αλληλουχίες (που ονομάστηκαν "αναγνώσματα") συναρμολογήθηκαν σε ένα συνεχές κείμενο με βάση τα επικαλυπτόμενα άκρα τους
Τα δείγματα καθαρισμένου DNA διαχωρίζονται σε μικρά θραύσματα, είτε με μηχανικές μεθόδους (π.χ. διατμητική διάτμηση με υπερήχους και νεφελοποίηση) είτε με ενζυματική πέψη.
Η ενίσχυση των προτύπων πραγματοποιείται με PCR σε στερεή επιφάνεια ή με PCR γαλακτώματος σε ξεχωριστούς μικροαντιδραστήρες, σφαιρίδια ή σφαίρες εντός των ακολουθιογράφων
Η μεροληψία στα δεδομένα NGS έχει παρατηρηθεί εκτενώς, αλλά δεν υπάρχει συμφωνία όσον αφορά τις πηγές της παρατηρούμενης μεροληψίας
Οι Benjamini και Speed υπέθεσαν ότι η σημαντικότερη αιτία της GC μεροληψίας ήταν η PCR
Η μεροληψία στις αναγνώσεις NGS συσχετίζεται στενά με την μεροληψία που παράγεται από τον ηχοβολισμό καθαρών τμημάτων DNA περιορισμού
Ανέφεραν και ανέλυσαν τη μεροληψία που παράγεται από τρεις μεθόδους διάτμησης του DNA, τον ηχοβολισμό, τον νεφελοποίηση και το Covaris και απέδειξαν ότι οι περιπτώσεις μεροληψίας στις διάφορες μεθόδους θραυσματοποίησης συσχετίζονται σε μεγάλο βαθμό μεταξύ τους και είναι κοινές για όλες τις υδροδυναμικές μεθόδους διάτμησης του DNA
Όλες οι υπάρχουσες τεχνολογίες αλληλούχισης δεν ανταποκρίνονται στο ιδανικό της ομοιόμορφης κατανομής των αναγνώσεων στο γονιδίωμα χωρίς διακυμάνσεις στην ποιότητα που εξαρτώνται από την αλληλουχία και αντίθετα παρουσιάζουν διάφορους τύπους και βαθμούς μεροληψίας.
Η μεροληψία της αλληλούχισης υποβαθμίζει τις εφαρμογές γονιδιωματικών δεδομένων, συμπεριλαμβανομένης της συναρμολόγησης γονιδιώματος και της ανακάλυψης παραλλαγών, οι οποίες βασίζονται στην κάλυψη σε όλο το γονιδίωμα.
Η μεροληψία κάλυψης είναι μια απόκλιση από την ομοιόμορφη κατανομή των αναγνώσεων σε όλο το γονιδίωμα.
Η μεροληψία σφάλματος είναι μια απόκλιση από την προσδοκία ομοιόμορφων ποσοστών αναντιστοιχιών, παρεμβολών και διαγραφών στις αναγνώσεις σε όλο το γονιδίωμα.
Οι τεχνολογίες αλληλούχισης είναι ευάλωτες σε πολλαπλές πηγές μεροληψίας:
Οι μέθοδοι που βασίζονται στη βακτηριακή κλωνοποίηση και τη χημική αλληλούχιση κατά Sanger υπόκεινται σε πολλές μεροληψίες που μειώνουν την κάλυψη, ιδίως στα ακραία GC, στα παλίνδρομα, στις ανεστραμμένες επαναλήψεις και στις αλληλουχίες που είναι τοξικές για τον βακτηριακό ξενιστή.
Η αλληλούχιση Illumina έχει αποδειχθεί ότι χάνει την κάλυψη σε περιοχές με υψηλό ή χαμηλό GC, φαινόμενο που παρατηρείται και σε άλλες τεχνολογίες "επόμενης γενιάς".
Η ενίσχυση με PCR κατά την κατασκευή της βιβλιοθήκης είναι γνωστή πηγή υποκάλυψης περιοχών με ακραίο GC και παρόμοιες μεροληψίες μπορεί επίσης να εισαχθούν κατά τη διάρκεια της PCR γέφυρας για την ενίσχυση συστάδων στην κυψελίδα ροής Illumina.
Τα ειδικά σφάλματα της Illumina μπορούν να οδηγήσουν σε μεροληψίες κάλυψης μειώνοντας την απόδοση του ευθυγραμμιστή.
Η Ion Torrent, όπως και η 454, χρησιμοποιεί χημεία χωρίς τερματιστή, η οποία μπορεί να περιορίζει την ικανότητά της για ακριβή αλληλουχία μεγάλων ομοιοπολυμερών και μπορεί επίσης να είναι ευαίσθητη στις μεροληψίες κάλυψης που εισάγονται από την PCR γαλακτώματος κατά την κατασκευή της βιβλιοθήκης.
Η Complete Genomics χρησιμοποιεί επίσης ενίσχυση μαζί με μια πολύπλοκη διαδικασία κατασκευής βιβλιοθήκης.
Οι περιορισμοί της επεξεργασίας σήματος και της κλήσης βάσεων μπορεί να οδηγήσουν σε υποαντιπροσώπευση ή αυξημένα ποσοστά σφάλματος σε ορισμένες θέσεις, όπως και η ανακριβής ευθυγράμμιση.
Μια ανακριβής αναφορά ή διαφορές δείγματος-αναφοράς μπορεί να προκαλέσουν διακυμάνσεις στην κάλυψη ή την ακρίβεια που μπορεί να διαγνωστούν λανθασμένα ως μεροληψία αλληλούχισης
Ο κατακερματισμός του DNA είναι απαραίτητο βήμα στην προετοιμασία του δείγματος για τις περισσότερες πλατφόρμες αλληλούχισης.
Είναι σημαντικό να επιλεγεί μια μέθοδος κατακερματισμού που παράγει αξιόπιστα θραύσματα κατάλληλου μεγέθους χωρίς να εισάγει μεροληψία όσον αφορά τις θέσεις κοπής ή τη σύνθεση των βάσεων των θραυσμάτων.
Οι κύριες μέθοδοι που χρησιμοποιούνται για τον κατακερματισμό περιλαμβάνουν:
Επεξεργασίες με βάση ένζυμα
Ακουστική διάτμηση
Ηχοβολισμός
Φυγοκεντρική διάτμηση
Σημειακή διάτμηση
Κοπή με βελόνα
Οι περισσότερες από αυτές τις μεθόδους περιλαμβάνουν περιορισμούς και δυσκολίες όπως:
Υψηλό κόστος
Κατακερματισμός
Ευρεία κατανομή μεγέθους θραυσμάτων
Μη αναστρέψιμη βλάβη στα θραύσματα DNA
Τα αποτελέσματα μπορεί να εξαρτώνται από τη συγκέντρωση και/ή να μην είναι ιδιαίτερα αναπαραγώγιμα
Μέχρι στιγμής έχει χρησιμοποιηθεί ο υπερηχοτονισμός για την παρασκευή DNA κατάλληλου μεγέθους, αλλά η μέθοδος αυτή συνδέεται με σημαντική απώλεια δείγματος DNA
Παρά την ευρεία χρήση, η επίδραση του ενζυμικού κατακερματισμού στις προκύπτουσες αλληλουχίες δεν έχει αξιολογηθεί προσεκτικά
Μια ανάλυση αποκάλυψε σημαντικά μεγαλύτερο αριθμό επαναλαμβανόμενων τεχνητών SNVs/indels στις βιβλιοθήκες που έχουν υποστεί επεξεργασία με ενδονουκλεάση σε σύγκριση με εκείνες που δημιουργήθηκαν με υπερηχογράφημα
Αυτά τα τεχνουργήματα χαρακτηρίζονταν από παλίνδρομη δομή στο γονιδιωματικό πλαίσιο, μεροληψία θέσης στις αλληλουχίες και υποκαταστάσεις πολλών νουκλεοτιδίων.
Η αλληλούχιση του γονιδιώματος με τη χρήση σύλληψης υβριδισμού περιλαμβάνει πολλαπλά βήματα, όπως:
Επεξεργασία ιστού
Αποθήκευση ιστού
Απομόνωση DNA,
Κατακερματισμός του DNA
Υβριδοποίηση ανιχνευτή
Ενίσχυση βιβλιοθήκης
Αλληλούχιση
Ανάλυση πληροφορικής
Σφάλματα αλληλούχισης μπορούν να εισαχθούν σε οποιοδήποτε από αυτά τα στάδια και τα νουκλεοτίδια μπορούν να τροποποιηθούν περαιτέρω μέσω οξείδωσης κατά την επεξεργασία ιστών, την αποθήκευση ιστών, την απομόνωση του DNA και τον κατακερματισμό του DNA.
Τα σφάλματα ενσωμάτωσης νουκλεοτιδίων μπορούν με τη σειρά τους να δημιουργήσουν μεροληψίες στην αντίδραση πολυμεράσης, να επηρεάσουν την ακριβή ενίσχυση της βιβλιοθήκης και να δημιουργήσουν θόρυβο αλληλούχισης
Αν και έχουν καταβληθεί σημαντικές προσπάθειες για την ελαχιστοποίηση αυτού του θορύβου αλληλούχισης πειραματικά, τα στοχαστικά σφάλματα παραμένουν επίμονα
Η φυσική διασπορά του διαλύματος DNA κατά τη διαδικασία οδηγεί συχνά σε απώλεια δείγματος DNA
Διατίθενται διάφορα εμπορικά κιτ προετοιμασίας βιβλιοθήκης, ωστόσο, αν και τα κιτ αυτά ελαχιστοποιούν την απώλεια DNA, παραμένει σε μεγάλο βαθμό άγνωστο σε ποιο βαθμό τα σφάλματα αλληλούχισης προκαλούνται από τη διαδικασία ενζυμικού κατακερματισμού
Η ποιότητα της αλληλούχισης εξαρτάται τόσο από την τυχαιότητα του κατακερματισμού του DNA όσο και από την επικάλυψη των θραυσμάτων που προκύπτουν
Επειδή το μέγεθος των θραυσμάτων τείνει να διαφέρει μεταξύ των πλατφορμών NGS και των τμημάτων αλληλούχισης, ο αποτελεσματικός έλεγχος του μεγέθους των θραυσμάτων DNA είναι επιτακτικός.
Εικάζεται ότι οι μεταλλάξεις προκύπτουν μετά τον ενζυμικό κατακερματισμό κατά τη διάρκεια της "διαδικασίας συμπλήρωσης" που ενορχηστρώνεται από την πολυμεράση του DNA για την επιδιόρθωση του άκρου
Ο υπερηχητικός συντονισμός διασπά τυχαία μόρια DNA σε διαφορετικές γονιδιωματικές θέσεις και, ως εκ τούτου, κατά την επακόλουθη διαδικασία συμπλήρωσης, νουκλεοτίδια ενσωματώνονται σε διαφορετικές γονιδιωματικές θέσεις σε διαφορετικά μόρια DNA.
Επειδή η ενδονουκλεάση HyperPlus κόβει κατά προτίμηση συγκεκριμένες θέσεις στο DNA, όταν ενσωματώνεται ένα λανθασμένο νουκλεοτίδιο, το προκύπτον τεχνούργημα θα μπορούσε λανθασμένα να αναγνωριστεί ως μετάλλαξη επειδή εμφανίζεται επανειλημμένα στην ίδια θέση σε διαφορετικά μόρια.
Θεωρήθηκε ότι ο θόρυβος θα μπορούσε να αποφευχθεί με τη χρήση της ίδιας μεθόδου κατακερματισμού του DNA για ζευγαρωμένα δείγματα, ωστόσο, παρόλο που υπήρξε σημαντική μείωση του αριθμού των SNVs/indels με τη χρήση μόνο μιας μεθόδου κατακερματισμού, κατά την προσεκτική εξέταση, εντοπίστηκε επιμονή του θορύβου HyperPlus μεταξύ των SNVs/indels που προέκυψαν από το συνδυασμό HH.
Αυτός ο θόρυβος ταξινομήθηκε συχνά από τον αλγόριθμο σε άλλες συγκρίσεις ανά ζεύγη και χαρακτηρίστηκε από παλίνδρομη δομή
Το εύρημα αυτό ενίσχυσε την πρόταση για τον κίνδυνο τα επίμονα σφάλματα να συγχέονται με γνήσιες μεταλλάξεις λόγω της επαναλαμβανόμενης εμφάνισής τους σε μια κοόρτη
Για τα γονιδιώματα RNA, η ροή εργασίας του RNA-seq είναι εξαιρετικά περίπλοκη και είναι εύκολο να παραχθεί μεροληψία
Αυτό μπορεί να βλάψει την ποιότητα του συνόλου δεδομένων RNA-seq και να οδηγήσει σε εσφαλμένη ερμηνεία του αποτελέσματος της αλληλούχισης
Το RNA-seq είναι μια εξαιρετικά περίπλοκη διαδικασία που περιλαμβάνει:
εκχύλιση και καθαρισμό RNA
κατασκευή βιβλιοθήκης (συμπεριλαμβανομένου του κατακερματισμού)
Γραμμική ενίσχυση ή ενίσχυση με PCR
Αλληλούχιση
Βιοπληροφορική ανάλυση
Αυτές οι διαδικασίες μπορούν αναπόφευκτα να εισάγουν ορισμένες αποκλίσεις που επηρεάζουν την ποιότητα των συνόλων δεδομένων RNA-seq και έχουν ως αποτέλεσμα τη λανθασμένη ερμηνεία τους.
Επομένως, η κατανόηση αυτών των μεροληψιών είναι ζωτικής σημασίας για την αποφυγή εσφαλμένης ερμηνείας των δεδομένων και για την αξιοποίηση του πλήρους δυναμικού αυτής της τεχνολογίας.
Το πιο προβληματικό βήμα στις διαδικασίες προετοιμασίας δειγμάτων είναι η ενίσχυση λόγω του γεγονότος ότι η ενίσχυση PCR εισάγει στοχαστικά μεροληψίες, οι οποίες μπορούν να μεταδοθούν σε μεταγενέστερους κύκλους
Επιπλέον, η PCR ενισχύει επίσης διαφορετικά μόρια με άνισες πιθανότητες, οδηγώντας στην άνιση ενίσχυση των μορίων cDNA
Το RNA είναι συνήθως κατακερματισμένο λόγω του περιορισμού του μήκους ανάγνωσης (<600 bp) των τεχνολογιών αλληλούχισης και της ευαισθησίας της ενίσχυσης σε μεγάλα μόρια cDNA
Υπάρχουν δύο κύριες προσεγγίσεις κατακερματισμού του RNA: η χημική (με χρήση ιόντων μετάλλων) και η ενζυμική (με χρήση RNase III)
Συνήθως, το RNA κατακερματίζεται με τη χρήση ιόντων μετάλλων, όπως Mg++ και Zn++, σε υψηλές θερμοκρασίες και αλκαλικές συνθήκες
Ωστόσο, η μέθοδος της τρανσποζάσης Tn5 έδειξε μεροληψία ανάλογα με την αλληλουχία, η οποία είναι η προτιμώμενη μέθοδος όταν είναι διαθέσιμες μόνο μικρές ποσότητες cDNA
Η φυσική μέθοδος περιλαμβάνει την ακουστική διάτμηση, τον ηχητικό καθαρισμό και την υδροδυναμική, οι οποίες μπορούν επίσης να παρουσιάσουν μη τυχαία μεροληψία κατακερματισμού του DNA
Όλες οι μελέτες αλληλούχισης περιορίζονται από την ακρίβεια των υποκείμενων πειραμάτων αλληλούχισης, επειδή η τεχνολογία RNA-seq μπορεί να εισάγει διάφορα σφάλματα και μεροληψίες κατά την προετοιμασία του δείγματος, την κατασκευή της βιβλιοθήκης, την αλληλούχιση και την απεικόνιση κ.λπ.
Τόσο τα νουκλεϊκά οξέα όσο και τα μεταλλικά ιόντα παρουσιάζουν σημαντική πολυπλοκότητα στις αλληλεπιδράσεις τους και οι αλληλεπιδράσεις αυτές θα μπορούσαν να επηρεάσουν τις χημικές και βιοχημικές ιδιότητες και των δύο μερών.
Τα περισσότερα πολυδύναμα ιόντα μετάλλων (M22+) μπορούν να προκαλέσουν διάσπαση (δηλαδή θραύση, διάσπαση, κατακερματισμό, αποπολυμερισμό ή ρήξη) των νουκλεϊκών οξέων.
Η εξειδίκευση αυτών των αντιδράσεων εξαρτάται τόσο από τη διαμόρφωση του νουκλεϊκού οξέος και τους τρόπους πρόσδεσης του μετάλλου όσο και από τις ιδιότητες των μεταλλικών ιόντων
Υπάρχουν διάφοροι παράγοντες που συχνά αγνοούνται και οι οποίοι μπορούν να έχουν δραματικές επιπτώσεις στα πειράματα ή μπορεί να οδηγήσουν σε τεχνουργήματα ή σε παρερμηνεία των πειραματικών δεδομένων
Η μόλυνση των διαλυμάτων από RNάσες και ιόντα Μ2+ μπορεί να οδηγήσει σε ταχεία αποικοδόμηση του RNA ακόμη και σε ουδέτερο pH και χαμηλές θερμοκρασίες
Η λήψη ακριβών μετρήσεων ποσοτικοποίησης μεταγραφών από το RNA-Seq έχει αποδειχθεί δύσκολη
Ένας από τους κύριους λόγους της ανακρίβειας είναι η αποτυχία των στατιστικών μοντέλων που χρησιμοποιούνται για την εξαγωγή των μετρήσεων να αναπαραστήσουν σωστά τις μεροληψίες που είναι εγγενείς στα δεδομένα RNA-Seq
Το στατιστικό μοντέλο της αρχικής έκδοσης του Cufflinks, για παράδειγμα, υποθέτει ότι τα θραύσματα cDNA που παράγονται από το RNA-Seq κατανέμονται ομοιόμορφα κατά μήκος του μεταγράφου
Στην πραγματικότητα, ωστόσο, η υπόθεση αυτή σπάνια πληρούται και, ως εκ τούτου, οι μετρήσεις ποσοτικοποίησης με αυτή την έκδοση του Cufflinks είναι συχνά ανακριβείς
Ένας τύπος μεροληψίας που επηρεάζει την ποσοτικοποίηση μεταγραφών από δεδομένα RNA-Seq είναι το αποτέλεσμα της προτίμησης του κατακερματισμού, δηλαδή της διαδικασίας που παράγει θραύσματα cDNA από μεταγραφή RNA, να παράγει θραύσματα σε ορισμένες θέσεις εντός της μεταγραφής, π.χ. στην αρχή ή/και στο τέλος της μεταγραφής.
Η μεροληψία θέσης μπορεί επίσης να προκληθεί από μεροληψία στο ίδιο το RNA, για παράδειγμα, λόγω αποικοδόμησης του RNA που έχει ως αποτέλεσμα τη συντόμευση του RNA.
Ένα άλλο είδος μεροληψίας στην RNA-Seq είναι η ειδική μεροληψία ακολουθίας που εισάγεται κατά τη διάρκεια της σύνδεσης, της ενίσχυσης και της αλληλούχισης NGS
Οι σημερινοί τεχνολογικοί περιορισμοί των ακολουθιογράφων απαιτούν τα μόρια cDNA να αντιπροσωπεύουν μόνο μερικά θραύσματα του RNA που εξετάζεται
Η τυχαιότητα που ενυπάρχει σε πολλά από τα στάδια προετοιμασίας για RNA-Seq οδηγεί σε θραύσματα των οποίων τα σημεία εκκίνησης (σε σχέση με τα μεταγράμματα από τα οποία αλληλουχήθηκαν) φαίνεται να επιλέγονται περίπου ομοιόμορφα τυχαία
Η παρατήρηση αυτή αποτέλεσε τη βάση των παραδοχών που διέπουν ορισμένες προσεγγίσεις ανάλυσης RNA-Seq οι οποίες, με όρους επιστήμης των υπολογιστών, αντιστρέφουν την "αναγωγή" της εκτίμησης του μεταγραφώματος στην αλληλούχιση DNA
Πρόσφατη προσεκτική ανάλυση αποκάλυψε μεροληψίες τόσο ως προς τη θέση όσο και ως προς την αλληλουχία στα αλληλουχθέντα θραύσματα
Αυτές οι μεροληψίες μπορούν να επηρεάσουν τις εκτιμήσεις έκφρασης και, ως εκ τούτου, είναι σημαντικό να διορθώνονται κατά τη διάρκεια της ανάλυσης RNA-Seq
Παρόλο που πολλές μεροληψίες μπορούν να αποδοθούν σε ιδιαιτερότητες των πρωτοκόλλων παρασκευής, δεν είναι επί του παρόντος δυνατό να προβλεφθεί η κατανομή των θραυσμάτων απευθείας από ένα πρωτόκολλο.
Αυτό οφείλεται σε πολλούς παράγοντες, συμπεριλαμβανομένης της αβεβαιότητας στη βιοχημεία πολλών βημάτων και του άγνωστου σχήματος και της επίδρασης της δευτερογενούς δομής του RNA σε ορισμένες διαδικασίες
Θα πρέπει να είναι προφανές ότι ανεξάρτητα από τη μέθοδο που χρησιμοποιείται για τον κατακερματισμό του DNA/RNA, οι μεροληψίες/σφάλματα είναι αναπόφευκτο φαινόμενο. Αυτό οδηγεί σε πολλές ανακρίβειες και στην αδυναμία αναπαραγωγής των αποτελεσμάτων. Όσο περισσότερα βήματα απαιτούνται για να χειραγωγηθεί και να διαμορφωθεί το δείγμα σε κάτι που μπορεί να διαβάσει η περιορισμένη τεχνολογία, τόσο πιο μακριά από την πραγματικότητα βρίσκεται το δείγμα. Η εμφάνιση μεροληψιών, σφαλμάτων και τεχνουργημάτων κατά τον κατακερματισμό μπορεί να ξεκινήσει μια αλυσιδωτή αντίδραση που θα καταστρέψει εντελώς κάθε προσπάθεια αλληλούχισης. Αυτό δεν λαμβάνει καν υπόψη τη μόλυνση και την πιθανή υποβάθμιση του RNA που είναι βέβαιο ότι θα συμβεί κατά τις διαδικασίες εκχύλισης που γίνονται στο δείγμα πριν από τον κατακερματισμό. Εάν τα πρώτα βήματα που εμπλέκονται στη δημιουργία ενός γονιδιώματος είναι γεμάτα με τόσο μεγάλες πιθανότητες παρεμβολής στη διαδικασία αλληλούχησης που τελικά οδηγούν σε αναξιόπιστα αποτελέσματα, ποια πίστη μπορεί να υπάρχει στο τελικό προϊόν ως προς την ακριβή αναπαράσταση;
Αν σας άρεσε αυτό το άρθρο και θα θέλατε να βοηθήσετε να στηρίξετε το συνεχές έργο μου, ο παρακάτω σύνδεσμος είναι μια επιλογή.
Παρακαλώ βοηθήστε να στηρίξετε το έργο μου. 🙏


ΦΑΚΕΛΛΟΣ "ΙΟΛΟΓΙΑ": Αλήθεια, Πόσο Ακριβή και Αξιόπιστα Είναι τα Γονιδιώματα;

Μετάφραση: Απολλόδωρος 22 Ιανουαρίου 2022 | MIKE STONE | Διαβάστε το εδώ
----Δικτυογραφία :
DNA and RNA Shearing/Fragmentation – ViroLIEgy
https://viroliegy.com/2022/02/15/dna-and-rna-shearing-fragmentation/